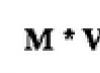Периферическая нервная система–комплекс анатомических образований, осуществляющих связь центральной нервной системы с кожным покровом, опорно-двигательным аппаратом, внутренними органами.
Развитие:
в начале 1-го месяца эмбрионального развития происходит образование нервной пластинки, при замыкании которой в нервную трубку выделяются зачатки межпозвонковых спинномозговых ганглиев и зачатки околопозвоночных узлов симпатического ствола. При этом клетки зачатков симпатической части автономной нервной системы начинают мигрировать в направлении ближайшего отрезка брюшного корешка, формируя соединительные ветви. В дальнейшем путем миграции нейробластов и роста отростков образуются предпозвоночные и интрамуральные сплетения автономной нервной системы.
В нервной трубке различные ее части растут неравномерно, что приводит к выделению основных отделов будущего спинного мозга: боковые стенки идут на построение серого вещества, а вентральные и дорсальные части - вентральных и дорсальных рогов. Зачатки спинного мозга образованы клетками двух родов: одни - спонгиобласты - образуют нейроглию, другие - нейробласты - развиваются в нейроциты.
На 3 - 4-й неделе развития отростки нейробластов нервной трубки выходят из нее и образуют метамерно расположенные брюшные корешки спинного мозга. Нейробласты, лежащие в зачатках спинномозговых узлов, также отдают длинные отростки, которые формируют спинные корешки. На 5 - 6-й неделе развития совершается слияние брюшного и спинного корешков с образованием смешанных спинномозговых нервов и их основных ветвей (брюшной, спинной, соединительной, оболочечной).
На 2-м месяце развития дифференцируются зачатки конечностей, в которые врастают нервные волокна соответствующих закладке сегментов. В первой половине 2-го месяца в связи с перемещением метамеров, формирующих конечности, образуются нервные сплетения. У человеческого эмбриона длиной 10 мм хорошо заметно плечевое сплетение, представляющее собой пластинку из отростков нервных клеток и нейроглии, которая на уровне проксимального конца развивающегося плеча делится на две: дорсальную и вентральную. Из дорсальной пластинки формируется в дальнейшем задний пучок, дающий начало подкрыльцовому и лучевому нервам, а из передней - боковой и медиальный пучки сплетения.
У эмбриона длиной 15 - 20 мм все нервные стволы конечностей и туловища соответствуют положению нервов у новорожденного. При этом формирование нервов туловища и нервов нижней конечности совершается подобным же путем, но несколько позже (на 2 недели).
Сравнительно рано (у эмбрионов длиной 8 - 10 мм) наблюдается проникновение мезенхимных клеток вместе с кровеносными сосудами. Мезенхимные клетки делятся и образуют внутриствольные оболочки нерва: эндо-, пери- и эпиневрий. Глиальные элементы зачатков спонгиобластов идут на построение шванновских оболочек длинных отростков нервных клеток. Миелинизация нервных волокон начинается неодновременно, с 3 - 4-го месяца эмбрионального развития, и заканчивается после рождения. Раньше миелинизируются черепные нервы, нервы верхних конечностей, позже - нервы туловища и нижних конечностей.
Состав:В ее состав входят сенсорные компоненты (сенсорные рецепторы и первичные афферентные нейроны) и двигательные компоненты (соматические мотонейроны и вегетативные мотонейроны).
Сенсорные рецепторы - структуры, воспринимающие воздействие разнообразных видов внешней энергии на организм. Они расположены на периферических окончаниях первичных афферентных нейронов, передающих получаемую рецепторами информацию в центральную нервную систему через посредство задних (дорсальных) корешков либо черепных нервов. Тела их клеток находятся в ганглиях задних корешков (спинномозговых, или спинальных ганглиях) либо в ганглиях черепных нервов. Ганглий периферической нервной системы - это скопление тел нейронов, выполняющих одинаковые функции.
К двигательному компоненту периферической нервной системы относятся соматические мотонейроны и вегетативные (автономные) мотонейроны. Тела соматических мотонейронов находятся в спинном мозгу или в стволе мозга. Они иннервируют волокна скелетных мышц. Как правило, у них длинные дендриты, получающие много синаптических входов. Мотонейроны каждой мышцы составляют определенное двигательное ядро. Ядро - это группа нейронов центральной нервной системы (ЦНС), имеющих одинаковые функции (не путать с ядром клетки). Например, от мотонейронов ядра лицевого нерва иннервируются мимические мышцы лица. Аксоны соматических мотонейронов покидают ЦНС через передний корешок либо через черепной нерв.
Вегетативные (автономные) мотонейроны посылают нервы к волокнам гладкой мускулатуры и к железам. Эти мотонейроны - вегетативные преганглионарные нейроны и вегетативные постганглионарные нейроны симпатической нервной системы и парасимпатической нервной системы.
Преганглионарные нейроны расположены в ЦНС - в спинном мозгу либо в стволе мозга. В отличие от соматических мотонейронов вегетативные преганглионарные нейроны образуют синапсы не прямо на своих эффекторных клетках (в гладкой мускулатуре или железах), а на постганглионарных нейронах, которые, в свою очередь, синаптически контактируют уже непосредственно с эффекторами.
Центральная нервная система анализирует сенсорную информацию, получаемую от сенсорных рецепторов, локализованных в окончаниях аксонов первичных афферентных нейронов. На основе этой информации она вырабатывает двигательные команды, которые передаются:
По двигательным аксонам от соматических мотонейронов к волокнам скелетных мышц;
Через вегетативные преганглионарные нейроны и вегетативные постганглионарные нейроны к миокарду, гладкой мускулатуре, железам. Таким образом ЦНС ощущает и анализирует окружающую среду, чтобы обеспечить адекватное поведение.
Аксоны первичных афферентных нейронов, соматических мотонейронов и вегетативных мотонейронов входят в состав переферической нервной системы (рис. 33.1). Таким образом, периферическая нервная система служит связующим звеном между ЦНС и окружающей средой.
Периферическая нервная система образована узлами (спинномозговыми, черепными и вегетативными), нервами (31 пара спинно-мозговых и 12 пар черепных) и нервными окончаниями, которые и обеспечивают связь ЦНС со всеми рецепторами и эффекторами организма.
В состав периферической нервной системы включают также черепные, спинно-мозговые и вегетативные ганглии, представляющие собой скопления тел нейронов за пределами ЦНС. Большинство периферических структур содержит чувствительные, двигательные и вегетативные волокна.
Нервная система человека подразделяется на центральную, периферическую и автономную части. Периферическая часть нервной системы представляет собой совокупность спинномозговых и черепных нервов. К ней относятся образуемые нервами ганглии и сплетения, а также чувствительные и двигательные окончания нервов. Таким образом, периферическая часть нервной системы объединяет все нервные образования, лежащие вне спинного и головного мозга. Такое объединение в известной мере условно, так как эфферентные волокна, входящие в состав периферических нервов, являются отростками нейронов, тела которых находятся в ядрах спинного и головного мозга. С функциональной точки зрения периферическая часть нервной системы состоит из проводников, соединяющих нервные центры с рецепторами и рабочими органами. Анатомия периферических нервов имеет большое значение для клиники, как основа для диагностики и лечения заболеваний и повреждений этого отдела нервной системы.
Строение нервов
Периферические нервы состоят из волокон, имеющих различное строение и неодинаковых в функциональном отношении. В зависимости от наличия или отсутствия миелиновой оболочки волокна бывают миелиновые (мякотные) или безмиелиновые (безмякотные). По диаметру миелиновые нервные волокна подразделяются на тонкие (1-4 мкм), средние (4-8 мкм) и толстые (более 8 мкм). Существует прямая зависимость между толщиной волокна и скоростью проведения нервных импульсов. В толстых миелиновых волокнах скорость проведения нервного импульса составляет примерно 80-120 м/с, в средних - 30-80 м/с, в тонких - 10-30 м/с. Толстые миелиновые волокна являются преимущественно двигательными и проводниками проприоцептивной чувствительности, средние по диаметру волокна проводят импульсы тактильной и температурной чувствительности, а тонкие - болевой. Безмиелиновые волокна имеют небольшой диаметр - 1-4 мкм и проводят импульсы со скоростью 1-2 м/с. Они являются эфферентными волокнами вегетативной нервной системы.
Таким образом, по составу волокон можно дать функциональную характеристику нерва. Среди нервов верхней конечности наибольшее содержание мелких и средних миелиновых и безмиелиновых волокон имеет срединный нерв, а наименьшее число их входит в состав лучевого нерва, локтевой нерв занимает в этом отношении среднее положение. Поэтому при повреждении срединного нерва бывают особенно выражены болевые ощущения и вегетативные расстройства (нарушения потоотделения, сосудистые изменения, трофические расстройства). Соотношение в нервах миелиновых и безмиелиновых, тонких и толстых волокон индивидуально изменчиво. Например, количество тонких и средних миелиновых волокон в срединном нерве может у разных людей колебаться от 11 до 45%.
Нервные волокна в стволе нерва имеют зигзагообразный (синусоидальный) ход, что предохраняет их от перерастяжения и создает резерв удлинения в 12-15% от их первоначальной длины в молодом возрасте и 7-8% в пожилом возрасте.
Нервы обладают системой собственных оболочек. Наружная оболочка, эпиневрий, покрывает нервный ствол снаружи, отграничивая его от окружающих тканей, и состоит из рыхлой неоформленной соединительной ткани. Рыхлая соединительная ткань эпиневрия выполняет все промежутки между отдельными пучками нервных волокон. Некоторые авторы называют эту соединительную ткань внутренним эпиневрием, в отличие от наружного эпиневрия, окружающего нервный ствол снаружи.
В эпиневрии в большом количестве находятся толстые пучки коллагеновых волокон, идущих преимущественно продольно, клетки фибробластического ряда, гистиоциты и жировые клетки. При изучении седалищного нерва человека и некоторых животных установлено, что эпиневрия состоит из продольных, косых и циркулярных коллагеновых волокон, имеющих зигзагообразный извилистый ход с периодом 37-41 мкм и амплитудой около 4 мкм. Следовательно, эпиневрия - очень динамичная структура, которая защищает нервные волокна при растяжении и изгибе.
Из эпиневрия выделен коллаген I типа, фибриллы которого имеют диаметр 70-85 нм. Однако некоторые авторы сообщают о выделении из зрительного нерва и других типов коллагена, в частности III, IV, V, VI. Нет единого мнения о природе эластических волокон эпиневрия. Одни авторы считают, что в эпиневрии отсутствуют зрелые эластические волокна, но обнаружены два вида близких к эластину волокон: окситалановые и элауниновые, которые располагаются параллельно оси нервного ствола. Другие исследователи считают их эластическими волокнами. Жировая ткань является составной частью эпиневрия. Седалищный нерв содержит обычно значительное количество жира и этим заметно отличается от нервов верхней конечности.
При исследовании черепных нервов и ветвей крестцового сплетения взрослых людей установлено, что толщина эпиневрия колеблется в пределах от 18-30 до 650 мкм, но чаще составляет 70-430 мкм.
Эпиневрий - в основном питающая оболочка. В эпиневрии проходят кровеносные и лимфатические сосуды, vasa nervorum, которые проникают отсюда в толщу нервного ствола.
Следующая оболочка, периневрий, покрывает пучки волокон, из которых состоит нерв. Она является механически наиболее прочной. При световой и электронной микроскопии установлено, что периневрий состоит из нескольких (7-15) слоев плоских клеток (периневрального эпителия, нейротелия) толщиной от 0.1 до 1.0 мкм, между которыми располагаются отдельные фибробласты и пучки коллагеновых волокон. Из периневрия выделен коллаген III типа, фибриллы которого имеют диаметр 50-60 нм. Тонкие пучки коллагеновых волокон расположены в периневрии без особого порядка. Тонкие коллагеновые волокна образуют в периневрии двойную спиральную систему. Причем волокна образуют в периневрии волнистые сети с периодичностью около 6 мкм. Установлено, что пучки коллагеновых волокон имею в периневрии плотное расположение и ориентированы как в продольном, так и концентрическом направлениях. В периневрии найдены элауниновые и окситалановые волокна, ориентированные преимущественно продольно, причем первые в основном локализуются в поверхностном его слое, а вторые - в глубоком слое.
Толщина периневрия в нервах с многопучковой структурой находится в прямой зависимости от величины покрываемого им пучка: вокруг мелких пучков не превышает 3-5 мкм, крупные пучки нервных волокон покрываются периневральным футляром толщиной от 12-16 до 34-70 мкм. Данные электронной микроскопии свидетельствуют, что периневрий имеет гофрированную, складчатую организацию. Периневрию придается большое значение в барьерной функции и обеспечении прочности нервов.
Периневрий, внедряясь в толщу нервного пучка, образует там соединительнотканные перегородки толщиной 0.5-6.0 мкм, которые делят пучок на части. Подобная сегментация пучков чаще наблюдается в поздних периодах онтогенеза.
Периневральные влагалища одного нерва соединяются с периневральными влагалищами соседних нервов, и через эти соединения происходит переход волокон из одного нерва в другой. Если учесть все эти связи, то периферическую нервную систему верхней или нижней конечности можно рассматривать как сложную систему связанных между собой периневральных трубок, по которым осуществляется переход и обмен нервных волокон как между пучками в пределах одного нерва, так и между соседними нервами.
Самая внутренняя оболочка, эндоневрий, покрывает тонким соединительнотканным футляром отдельные нервные волокна. Клетки и внеклеточные структуры эндоневрия вытянуты и ориентированы преимущественно по ходу нервных волокон. Количество эндоневрия внутри периневральных футляров по сравнению с массой нервных волокон невелико. Эндоневрий содержит коллаген III типа с фибриллами диаметром 30-65 нм. Мнения о наличии в эндоневрии эластических волокон весьма противоречивы. Одни авторы считают, что эндоневрий не содержит эластических волокон. Другие обнаружили в эндоневрии близкие по свойствам к эластическим окситалановые волокна с фибриллами диаметром 10-12.5 нм, ориентированные, главным образом, параллельно аксонам.
При электронно-микроскопическом исследовании нервов верхней конечности человека обнаружено, что отдельные пучки коллагеновых фибрилл инвагинированы в толщу шванновских клеток, содержащих помимо этого еще и немиелинизированные аксоны. Коллагеновые пучки могут быть полностью изолированы клеточной мембраной от основной массы эндоневрия или только могут частично внедряться в клетку, находясь в контакте с плазматической мембраной. Но каким бы ни было расположение коллагеновых пучков, фибриллы всегда находятся в межклеточном пространстве, и никогда не были замечены во внутриклеточном. Такой тесный контакт шванновских клеток и коллагеновых фибрилл, по мнению авторов, увеличивает сопротивление нервных волокон различным растягивающим деформациям и укрепляет комплекс «шванновская клетка - немиелинизированный аксон».
Известно, что нервные волокна сгруппированы в отдельные пучки различного калибра. У разных авторов существуют различные определения пучка нервных волокон в зависимости от позиции, с которой эти пучки рассматриваются: с точки зрения нейрохирургии и микрохирургии или с точки зрения морфологии. Классическим определением нервного пучка является группа нервных волокон, ограниченная от других образований нервного ствола периневральной оболочкой. И этим определением руководствуются при исследовании морфологи. Однако при микроскопическом исследовании нервов часто наблюдаются такие состояния, когда несколько групп нервных волокон, прилежащих друг к другу, имеют не только собственные периневральные оболочки, но и окружены общим периневрием. Эти группы нервных пучков часто бывают видны при макроскопическом исследовании поперечного среза нерва во время нейрохирургического вмешательства. И эти пучки чаще всего описываются при клинических исследованиях. Из-за различного понимания строения пучка происходят в литературе противоречия при описании внутриствольного строения одних и тех же нервов. В связи с этим ассоциации нервных пучков, окруженные общим периневрием, получили название первичных пучков, а более мелкие, их составляющие, - вторичных пучков.
На поперечном срезе нервов человека соединительнотканные оболочки (эпиневрий, периневрий) занимают значительно больше места (67.03-83.76%), чем пучки нервных волокон. Показано, что количество соединительной ткани зависит от числа пучков в нерве. Ее значительно больше в нервах с большим количеством мелких пучков, чем в нервах с немногими крупными пучками.
Показано, что пучки в нервных стволах могут располагаться относительно редко с промежутками в 170-250 мкм, и более часто - расстояние между пучками менее 85-170 мкм.
В зависимости от строения пучков выделяют две крайние формы нервов: малопучковую и многопучковую. Первая характеризуется небольшим количеством толстых пучков и слабым развитием связей между ними. Вторая состоит их множества тонких пучков с хорошо развитыми межпучковыми соединениями.
Когда количество пучков небольшое, пучки имеют значительные размеры, и наоборот. Малопучковые нервы отличаются сравнительно небольшой толщиной, наличием небольшого количества крупных пучков, слабым развитием межпучковых связей, частым расположением аксонов внутри пучков. Многопучковые нервы отличаются большей толщиной и состоят из большого количества мелких пучков, в них сильно развиты межпучковые связи, аксоны располагаются в эндоневрии рыхло.
Толщина нерва не отражает количества содержащихся в нем волокон, и не существует закономерностей расположения волокон на поперечном срезе нерва. Однако установлено, что в центре нерва пучки всегда тоньше, на периферии - наоборот. Толщина пучка не характеризует количества заключенных в нем волокон.
В строении нервов установлена четко выраженная асимметрия, то есть неодинаковое строение нервных стволов на правой и левой сторонах тела. Например, диафрагмальный нерв имеет слева большее количество пучков, чем справа, а блуждающий нерв - наоборот. У одного человека разница в количестве пучков между правым и левым срединными нервами может варьировать от 0 до 13, но чаще составляет 1-5 пучков. Разница в количестве пучков между срединными нервами разных людей равняется 14-29 и с возрастом увеличивается. В локтевом нерве у одного и того же человека разница между правой и левой сторонами в количестве пучков может колебаться от 0 до 12, но чаще составляет также 1-5 пучков. Различие в количестве пучков между нервами разных людей достигает 13-22.
Разница между отдельными субъектами в количестве нервных волокон колеблется в срединном нерве от 9442 до 21371, в локтевом нерве - от 9542 до 12228. У одного и того же человека разница между правой и левой стороной варьирует в срединном нерве от 99 до 5139, в локтевом нерве - от 90 до 4346 волокон.
Источниками кровоснабжения нервов являются соседние близлежащие артерии и их ветви. К нерву обычно подходят несколько артериальных ветвей, причем интервалы между входящими сосудами варьируют в крупных нервах от 2-3 до 6-7 см, а в седалищном нерве - до 7-9 см. Кроме того, такие крупные нервы, как срединный и седалищный, имеют собственные сопровождающие артерии. В нервах, имеющих большое количество пучков, в эпиневрии содержится много кровеносных сосудов, причем они имеют сравнительно малый калибр. Наоборот, в нервах с небольшим количеством пучков сосуды одиночные, но значительно более крупные. Артерии, питающие нерв, в эпиневрии Т-образно делятся на восходящую и нисходящую ветви. Внутри нервов артерии делятся до ветвей 6-го порядка. Сосуды всех порядков анастомозируют между собой, образуя внутриствольные сети. Эти сосуды играют значительную роль в развитии коллатерального кровообращения при выключении крупных артерий. Каждая артерия нерва сопровождается двумя венами.
Лимфатические сосуды нервов находятся в эпиневрии. В периневрии между его слоями образуются лимфатические щели, сообщающиеся с лимфатическими сосудами эпиневрия и эпиневральными лимфатическими щелями. Таким образом, по ходу нервов может распространяться инфекция. Из больших нервных стволов обычно выходят несколько лимфатических сосудов.
Оболочки нервов иннервируются ветвями, отходящими от данного нерва. Нервы нервов имеют в основном симпатическое происхождение и по функции являются сосудодвигательными.
Спинномозговые нервы
Развитие спинномозговых нервов
Развитие спинномозговых нервов связано как с развитием спинного мозга, так и формированием тех органов, которые иннервируют спинномозговые нервы.
В начале 1-го месяца внутриутробного развития у эмбриона по обеим сторонам нервной трубки закладываются нервные гребни, которые подразделяются, соответственно сегментам тела, на зачатки спинномозговых ганглиев. Нейробласты, находящиеся в них, дают начало чувствительным нейронам спинномозговых ганглиев. На 3-4-й неделе последние образуют отростки, периферические концы которых направляются к соответствующим дерматомам, а центральные концы врастают в спинной мозг, составляя задние (дорсальные) корешки. Нейробласты вентральных (передних) рогов спинного мозга посылают отростки к миотомам «своих» сегментов. На 5-6-й неделе развития в результате объединения волокон вентральных и дорсальных корешков формируется ствол спинномозгового нерва.
На 2-м месяце развития дифференцируются зачатки конечностей, в которые врастают нервные волокна соответствующих закладке сегментов. В 1-й половине 2-го месяца в связи с перемещением метамеров, формирующих конечности, образуются нервные сплетения. У человеческого эмбриона длиной 10 мм хорошо заметно плечевое сплетение, представляющее собой пластинку из отростков нервных клеток и нейроглии, которая на уровне проксимального конца развивающегося плеча делится на две: дорсальную и вентральную. Из дорсальной пластинки формируется в дальнейшем задний пучок, дающий начало подмышечному и лучевому нервам, а из передней - латеральный и медиальный пучки сплетения.
У эмбриона длиной 15-20 мм все нервные стволы конечностей и туловища соответствуют положению нервов у новорожденного. При этом формирование нервов туловища и нервов нижних конечностей совершается подобным же путем, но на 2 недели позже.
Сравнительно рано (у эмбриона длиной 8-10 мм) наблюдается проникновение в нервные стволы мезенхимных клеток вместе с кровеносными сосудами. Мезенхимные клетки делятся и образуют внутриствольные оболочки нервов. Миелинизация нервных волокон начинается с 3-4-го месяца эмбрионального развития и заканчивается на 2-м году жизни. Раньше миелинизируются нервы верхних конечностей, позже - нервы туловища и нижних конечностей.
Таким образом, каждая пара спинномозговых нервов осуществляет связь определенного сегмента спинного мозга с соответствующим сегментом тела зародыша. Эта связь сохраняется и в дальнейшем развитии зародыша. Сегментарная иннервация кожных покровов может быть выявлена у взрослого человека, она имеет большое значение в неврологической диагностике. Обнаружив расстройство чувствительности в том или ином участке тела, можно определить, какие сегменты спинного мозга затронуты патологическим процессом. Иначе обстоит дело с иннервацией мышц. Поскольку большинство крупных мышц образуется от слияния нескольких миотомов, каждая из них получает иннервацию из нескольких сегментов спинного мозга.
ПНС – отдел нервной системы, который совместно с центральной и вегетативной частью обеспечивает единство самой нервной ткани и всего организма. Главная функция ПНС обеспечить связь организма с внешним окружением за счет контролируемой высшими центрами рефлекторной деятельности.
Составляющие ПНС
К ПНС относят черепные нервы (ЧН). Важно помнить, ядра этих нервов имеют церебральную и спинальную (для добавочного нерва) локализацию, принадлежат они ПНС. Моторные нейроны, локализованные в передней части выступающего передние рога, также относятся к ПНС. Кроме этого, ПНС принадлежат чувствительные, двигательные и общие образующиеся при их слиянии корешки, ганглии, сплетения и собственно периферические нервы.Черепные нервы
Выделяют 12 пар ЧН. По факту все нервы смешанные, поэтому правильнее обозначать их как преимущественно двигательные или преимущественно чувствительные. Исключение составляют обонятельный и зрительный нервы (они чисто чувствительные).Первая пара – обонятельный нерв обеспечивает функцию обоняния.
Вторая пара – зрительный нерв проводит импульс от сетчатки глаза в затылочную кору. Первая и вторая пары являются редуцированными долями головного мозга, однако их относят к ПНС.
Третья, четвертая и шестая пары обеспечивают движения глаз (глазодвигательный, блоковый и отводящий).
Пятая пара – тройничный нерв дает чувствительность на половине лица, кроме того является двигательным для жевательных мышц.
Лицевой нерв двигательный для мимических мышц, кроме того в нем есть вегетативные порции, обеспечивающие иннервацию слезной железы и вкусовую чувствительность задней части языка.
Преддверно-улитковый нерв в своем названии объединяет два различных по функциям нерва. Преддверная часть проводит импульсы от вестибулярного аппарата, обеспечивая равновесие при перемене положения головы, улитковая - от кортиева органа и является слуховым нервом.
Языкоглоточный нерв совместно с блуждающим обеспечивают чувствительность мягкого неба, гортани, надгортанника, являются двигательными для мышц этих образований. Кроме того, блуждающий нерв является вегетативным парасимпатическим нервом.
Добавочный нерв анатомически является спинномозговым, так как его ядро заложено в верхних сегментах спинного мозга, а своим каудальным концом оно может доходить до шейного утолщения. Это двигательный нерв для трапецевидной, дельновидной и грудинно-ключично-сосцевидной мышц.
Спинномозговые нервы
Ядра нервных клеток спинномозговых клеток локализованы в передних рогах спинного мозга. Передний корешок является двигательным и обеспечивает моторику мышц. Задний корешок – чувствительный, собирает информацию о раздражении, нанесенном на кожу и слизистые. Они объединяются в общий спинномозговой корешок и составляют 31 пару.Периферические нервы
Являются смешанными, но можно выделить преимущественно чувствительные или двигательные порции. Периферический нерв представляет собой совокупность волокон, собранных в пучки с изоляцией соединительной ткани.Основные периферическе нервы, исходящие из плечевого сплетения:
подкрыльцовый нерв;
кожно-мышечный;
лучевой;
локтевой;
срединный;
кутанный внутренний нерв плеча и предплечья.
Основные периферическе нервы, исходящие из поясничного сплетения:
Бедренный;
Запирательный;
Наружный кутанный нерв бедра;
Бедренно-половой.
Основные периферическе нервы, исходящие из крестцового сплетения:
Седалищный;
Мало- и большеберцовый;
Верхне- и нижнеягодичный;
Задний кутанный нерв бедра.
Виды повреждения периферического нервного волокна следующие. При полной перерезке нерва, страдают осевой цилиндр и миелиновая оболочка волокна. Такое состояние называется Валлеровское перерождение нерва. Вторым вариантом повреждений является дегенерация самого аксона, при первичном поражении осевого цилиндра. Демиелинизация – частая причина развития ряда патологических состояний, затрагивающих ПНС, здесь поражена непосредственно миелиновая оболочка. Существует еще один вариант повреждения нервного волокна, когда поражено тело нервной клетки. Такой вид называют нейронопатией.
Периферическая нервная система (ПНС) – условно выделяемая часть нервной системы, структуры которой находятся вне головного и спинного мозга. К периферической нервной системе относятся 12 пар черепных нервов, их корешки, чувствительные и вегетативные ганглии, расположенные по ходу стволов и ветвей этих нервов, а также передние и задние корешки спинного мозга и 31 пара спинномозговых нервов, чувствительные ганглии, нервные сплетения, периферические нервные стволы туловища и конечностей, правый и левый симпатические стволы, вегетативные сплетения, ганглии и нервы. Каждый нерв состоит из нервных волокон, миелинизированных и немиелинизированных. Снаружи нерв окружен соединительно-тканой оболочкой – эпиневрием, в который входят питающие нерв сосуды. Нерв состоит из пучков, которые, в свою очередь, покрыты периневрием, а отдельные волокна – эндоневрием.
Условность анатомического разделения центральной и периферической нервной системы определяется тем, что нервные волокна, составляющие нерв, являются или аксонами двигательных нейронов, расположенных в передних рогах сегмента спинного мозга, или дендритами чувствительных нейронов межпозвоночных ганглиев (аксоны этих клеток направляются по задним корешкам в спинной мозг). Таким образом, тела нейронов расположены в центральной нервной системе, а их отростки – в периферической (для двигательных клеток), или, наоборот, отростки нейронов, расположенных в периферической нервной системе, составляют проводящие пути ЦНС (для чувствительных клеток).
Основная функция ПНС заключается в обеспечении связи ЦНС с внешней средой и органами-мишенями. Она осуществляется либо проведением нервных импульсов от экстеро-, проприо- и интерорецепторов к соответствующим сегментарным и надсегментарным образованиям спинного и головного мозга, либо в обратном направлении – регулирующих сигналов из ЦНС к мышцам, обеспечивающим перемещение тела в окружающем пространстве, к внутренним органам и системам.
В ПНС нервные стволы могут содержать двигательные волокна (передние корешки спинного мозга, лицевой, отводящий, блоковый, добавочный и подъязычный черепные нервы), чувствительные (задние корешки спинного мозга, чувствительная часть тройничного нерва, слуховой нерв) или вегетативные (висцеральные ветви симпатической и парасимпатической систем). Основная часть нервных стволов туловища и конечностей является смешанной (содержит двигательные, чувствительные и вегетативные волокна). К смешанным нервам относятся межреберные нервы, стволы шейного, плечевого и пояснично-крестцового сплетений и исходящих из них нервов верхних (лучевого, срединного, локтевого и др.) и нижних (бедренного, седалищного, большеберцового, глубокого малоберцового и др.) конечностей. Соотношение двигательных, чувствительных и вегетативных волокон в стволах смешанных нервов может значительно варьировать. Наибольшее количество вегетативных волокон содержат срединный и большеберцовый нервы, а также блуждающий нерв.
В строении ПНС имеется ряд закономерностей:
1) нервы являются парными и расходятся симметрично в стороны от головного и спинного мозга, лежащего по осевой линии тела;
2) нервы, подобно артериям, идут к органам по кратчайшему пути. Если в процессе внутриутробного развития орган перемещается, нерв, соответственно, удлиняется и следует за ним;
3) нервы, иннервирующие мышцы, отходят от тех сегментов спинного мозга, которые соответствуют миотомам, из которых происходят эти мышцы; при их последующем перемещении источник иннервации сохраняется вблизи зоны закладки;
4) нервные стволы сопровождают артерии, вены, лимфатические сосуды, образуя сосудисто-нервные пучки, располагающиеся на сгибательных поверхностях конечностей, будучи защищенными фасциальными влагалищами, мышцами.
Черепные нервы
От головного мозга отходят 12 пар черепных нервов, из них обонятельный и зрительный нервы являются, по существу, редуцированной частью мозга, остальные черепные нервы напоминают спинномозговые (рис. 8.21).
Обонятельный нерв (I пара ) – образован аксонами обонятельных рецепторных нейронов. Эти нейроны расположены в обонятельном эпителии, выстилающем верхнюю поверхность полости носа. Обонятельные нервные волокна собираются в 15–20 тонких обонятельных нервов. Нервы проходят через отверстия решетчатой пластинки черепа и заканчиваются в обонятельных луковицах головного мозга. Здесь находятся клетки второго нейрона. Аксоны этих клеток в составе обонятельных путей направляются к первичным (подкорковым) обонятельным центрам – в обонятельный треугольник, переднее продырявленное пространство, прозрачную перегородку, таламус. В этих образованиях находятся клетки третьего нейрона, аксоны которых направляются в височные доли своей и противоположной сторон, преимущественно в парагиппокампальную извилину. Таким образом, обонятельные импульсы из каждой половины носа поступают в оба полушария мозга.
Рис. 8.21. Топография выхода черепных нервов
Зрительный нерв (II пара) – начинается он в сетчатке – внутренней оболочке глаза. Здесь имеются рецепторы – палочки, воспринимающие черно-белое изображение, и колбочки, ответственные за цветное восприятие. Палочки и колбочки биполярными нейроцитами соединены с нейроцитами ганглиозного слоя сетчатки. Отростки ганглиозных нейронов собираются в пучок в области слепого пятна сетчатки глаза, образуя зрительный нерв. Он прободает сосудистую оболочку и склеру (внутриглазная часть нерва), проходит в глазнице (глазничная часть) к зрительному каналу черепа, проникает через него в полость черепа (внутриканальная часть) и сближается с парным нервом другой стороны.
На основании мозга, кпереди от турецкого седла, зрительные нервы образуют зрительный перекрест. Перекрещиваются только волокна, идущие от внутренних половин сетчатки обоих глаз. Позади зрительного перекреста образуются зрительные тракты, каждый из которых включает волокна от одноименных половин сетчатки обоих глаз. Так, в правый зрительный тракт попадают волокна от правых половин сетчатки, а в левый – от левых половин. Зрительные тракты заканчиваются в первичных (подкорковых) зрительных центрах – верхних холмиках, таламусе и наружных коленчатых телах. Волокна, идущие в верхние холмики, связывают зрительный нерв с глазодвигательным. В наружных коленчатых телах и таламусе располагаются клетки, аксоны которых направляются на внутреннюю поверхность затылочных долей, оканчиваясь в коре большого мозга по обе стороны шпорной борозды. Кверху от нее оканчиваются волокна, связывающие с корой верхние половины сетчатки, а книзу – волокна от нижних половин сетчатки.
Глазодвигательный нерв (III пара) – ядра его располагаются в ножках мозга под водопроводом среднего мозга. Через верхнюю глазничную щель нерв попадает в глазницу, где иннервирует следующие мышцы: поднимающую верхнее веко, верхнюю прямую (поворачивает глазное яблоко кверху), внутреннюю прямую (двигает глазное яблоко внутри), нижнюю прямую (двигает глазное яблоко книзу) и нижнюю косую (двигает глазное яблоко вверх и кнаружи). Глазодвигательный нерв имеет также вегетативные (парасимпатические) ядра. Выходящие из них волокна образуют в пределах глазницы глазодвигательный корешок, который прерывается в ресничном узле и иннервирует ресничную мышцу, изменяющую кривизну хрусталика, и мышцу, суживающую зрачок (сфинктер зрачка). Путем изменения кривизны хрусталика глаз приспосабливается к видению предметов на близком и далеком расстояниях (аккомодация). Сфинктер зрачка играет защитную роль: при освещении глаза зрачок суживается, и на сетчатку попадает меньше света. Зрачковый рефлекс осуществляется при участии волокон зрительного нерва, направляющихся к верхним холмикам (афферентная часть дуги), волокон, соединяющих верхние холмики с ядрами глазодвигательного нерва (вставочный нейрон), и парасимпатических волокон глазодвигательного нерва (эфферентная часть рефлекторной дуги).
Блоковой нерв (IV пара) – ядро этого нерва расположено в покрышке среднего мозга, на уровне нижних холмиков. Нерв выходит из ствола мозга через крышу среднего мозга и через верхнюю глазничную щель попадает в глазницу, где иннервирует верхнюю косую мышцу, поворачивающую глазное яблоко вниз и кнаружи.
Тройничный нерв (V пара) – нерв смешанный, осуществляет чувствительную и двигательную иннервацию. Ядра тройничного нерва расположены в стволе мозга. Состоит из трёх ветвей: глазничный нерв, верхнечелюстной и нижнечелюстной нерв. Из них первые две ветви чувствительные, третья содержит одновременно чувствительные и двигательные волокна.
Глазной нерв обеспечивает чувствительную иннервацию кожи лба, переднего отдела волосистой части головы, верхнего века, внутреннего угла глаза, спинки носа, конъюнктивы, верхней части слизистой оболочки носа и секреторную иннервацию слезной железы.
Верхнечелюстной нерв выходит из полости черепа через круглое отверстие, иннервирует кожу нижнего века, боковой поверхности носа, щек и верхней губы, нижние отделы слизистой оболочки носа, верхнюю челюсть и ее зубы.
Нижнечелюстной нерв выходит из полости черепа через овальное отверстие, иннервирует кожу нижней челюсти, щек, нижней губы, подбородка, нижнюю челюсть и ее зубы, слизистую оболочку щек, нижних отделов полости рта, языка, а также обеспечивает секреторную иннервацию подъязычной и поднижнечелюстной слюнных желез.
Отводящий нерв (IV пара) – двигательный. Ядро находится в области моста. Далее нерв через верхнюю глазничную щель направляется в глазницу и иннервирует латеральную прямую мышцу глазного яблока, которая поворачивает его кнаружи.
Отводящий нерв, глазодвигательный и блоковый составляют группу нервов, обеспечивающих подвижность глазных яблок. Синхронные повороты глаз в одном направлении совершаются благодаря связям между отдельными ядрами этой группы нервов. Такую связь обеспечивает задний продольный пучок, начинающийся в среднем мозге в ядрах заднего продольного пучка. Для осуществления сознательных движений глаз к заднему продольному пучку приходят импульсы из задних отделов средней лобной извилины противоположного полушария большого мозга.
Лицевой (промежуточно-лицевой) нерв (VII пара) – имеет ядра, расположенные в мосту. Нерв выходит на основание мозга в мостомозжечковом треугольнике. Затем нервные пучки через внутреннее слуховое отверстие входят в лицевой канал височной кости, в котором на уровне коленца разделяются на лицевой и промежуточный нервы. Промежуточный нерв включает чувствительные волокна, несущие вкусовые импульсы от передних двух третей языка, и парасимпатические волокна – к поднижнечелюстной и подъязычной слюнным железам, к слезной железе и железам слизистой оболочки полости носа и мягкого неба. Лицевой нерв выходит из черепа через шилососцевидное отверстие, образует сплетение в околоушной слюнной железе и разделяется на отдельные ветви, иннервирующие мышцы лица (носовые, скуловые, круговые мышцы глаза и рта и др.), а также некоторые мышцы шеи.
Преддверно-улитковый нерв (VIII пара). Выходит из черепа в виде улиткового и преддверного корешков. Улитковая часть нерва (собственно слуховой нерв) образуется в чувствительном улитковом (спиральном) узле, который находится во внутреннем ухе, в области улиткового лабиринта. Аксоны клеток улиткового узла образуют улитковый (слуховой) корешок, который через внутреннее слуховое отверстие попадает в полость черепа и оканчивается в улитковых ядрах, расположенных в задней части моста. Волокна клеток второго нейрона слухового пути, начавшись в улитковых ядрах и ядрах трапециевидного тела, направляются к первичным слуховым центрам – нижним холмикам и медиальным коленчатым телам. От медиальных коленчатых тел, где располагаются клетки третьего нейрона слухового пути, аксоны направляются в кору височных долей большого мозга. Благодаря частичному перекресту волокон вторых нейронов слуховые сигналы из каждого уха направляются в обе височные доли.
Преддверная (вестибулярная) часть нерва начинается в преддверном (чувствительном) узле, расположенном на дне внутреннего слухового прохода. Дендриты клеток этого узла направляются к рецепторным клеткам полукружных протоков внутреннего уха. Протоки частично заполнены жидкостью, которая раздражает рецепторы, перемещаясь при изменении положения тела или головы. Аксоны клеток преддверного узла образуют преддверный корешок, который направляется к преддверным ядрам моста (второй нейрон). Из таламуса идут волокна к коре височной доли большого мозга, где располагается корковое ядро вестибулярного анализатора. Кроме того, ядра мозжечка (ядро шатра) связаны с ядрами экстрапирамидной системы (мозжечково-красноядерный путь). Основной функцией преддверной части является сохранение равновесия. В связи с многочисленными связями ядер преддверной части нерва она участвует в регуляции непроизвольных двигательных актов.
Языкоглоточный нерв (IX пара ) – смешанный, имеет несколько ядер, расположенных в продолговатом мозге. Языкоглоточный нерв обеспечивает чувствительную иннервацию слизистой оболочки верхней части глотки, мягкого неба, задней трети языка и двигательную иннервацию мышц глотки, участвуя в глотании и артикуляции. Секреторные (парасимпатические) волокна языкоглоточного нерва оканчиваются в околоушной слюнной железе. Вкусовые волокна иннервируют заднюю треть языка.
Блуждающий нерв (X пара) – смешанный, имеет двигательное и чувствительное ядра и вегетативное (парасимпатическое) ядро. Блуждающий нерв – наиболее длинный из всех черепных нервов, так как сфера его иннервации распространяется от твердой оболочки головного мозга до сигмовидной ободочной кишки.
Значение блуждающего нерва очень велико, так как он обеспечивает чувствительную и вегетативную (парасимпатическую) иннервацию всех внутренних органов, кроме органов малого таза; значительную часть пищевого канала (до сигмовидной оболочной кишки), сердце, мышечную оболочку сосудов, трахею и легкие, железы слизистой оболочки пищевода, желудка и кишок, печень, поджелудочную железу, почки. Чувствительные волокна блуждающего нерва иннервируют также некоторые отделы твердой оболочки головного мозга и наружный слуховой проход с ушной раковиной. Двигательные волокна блуждающего нерва обеспечивают произвольные движения мышц глотки, мягкого неба и гортани. Таким образом, блуждающий нерв осуществляет нервную регуляцию таких жизненно важных функций, как дыхание и сердечно-сосудистая деятельность, а также участвует в осуществлении актов глотания и фонации.
Добавочный нерв (XI пара) – двигательный, иннервирует грудино-ключично-сосцевидную и трапециевидную мышцы.
Подъязычный нерв (XII пара) – двигательный, иннервирует мышцы языка.
Спинномозговые нервы
Спинномозговые нервы представляют собой парные нервные стволы, которые созданы слиянием двух корешков спинного мозга – заднего (чувствительного) и переднего (двигательного) (рис. 8.22).
На уровне межпозвоночного отверстия они соединяются, выходят и делятся на четыре ветви:
1) передние, иннервирующие кожу и мышцы конечностей и передней поверхности туловища;
2) задние, иннервирующие кожу и мышцы задней поверхности туловища; 3) менингеальные, направляющиеся к твердой оболочке спинного мозга;
4) соединительные, содержащие симпатические преганглионарные волокна, следующие к симпатическим узлам. Передние ветви спинномозговых нервов образуют сплетения: шейное, плечевое, пояснично-крестцовое и копчиковое.

Рис. 8.22. Схема образования спинномозгового нерва:
1 – ствол спинномозгового нерва; 2 – передний (двигательный) корешок; 3 – задний (чувствительный) корешок; 4 – корешковые нити; 5 – спинномозговой (чувствительный) узел; 6 –медиальная часть задней ветви; 7 – латеральная часть задней ветви; 8 – задняя ветвь; 9 – передняя ветвь; 10 – белая ветвь; 11 – серая ветвь; 12 – менингеальная ветвь
У человека находится 31 пара спинномозговых нервов, которые соответствуют 31 паре сегментов спинного мозга (8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 пара копчиковых нервов). Каждая пара спинномозговых нервов иннервирует определенный участок мышц (миотом), кожи (дерматом) и костей (склеротом). На основании этого выделяют сегментарную иннервацию мышц, кожи и костей.
Шейное сплетение образуется передними ветвями I – IV шейных нервов; иннервирует кожу затылка, боковой поверхности лица, над-, подключичную и верхнелопаточную области, диафрагму.
Плечевое сплетение образовано передними ветвями V, VI, VII, VIII и частично I грудного спинномозговых нервов (рис. 8.23).
Отделы нервной системы
Анатомическое подразделение отделов нервной системы:
(1) центральная нервная система (ЦНС) -
включает головной и спинной мозг;
(2) периферическая нервная система - включает периферические нервные ганглии (узлы), нервы и нервные окончания (описаны в разделе «Нервная ткань»).
Физиологическое подразделение отделов нервной системы (в зависимости от характера иннервации органов и тканей):
(1) соматическая (анимальная) нервная система - контролирует преимущественно функции произвольного движения;
(2) автономная (вегетативная) нервная система - регулирует деятельность внутренних органов, сосудов и желез.
Автономная нервная система подразделяется на взаимодействующие друг с другом симпатический и парасимпатический отделы, которые различаются локализацией периферических узлов и центров в мозгу, а также характером влияния на внутренние органы.
В соматическую и автономную нервную систему входят звенья, расположенные в ЦНС и периферической нервной системе. Функционально ведущей тканью органов нервной системы является нервная ткань, включающая нейроны и глию. Скопления нейронов в ЦНС обычно называют ядрами, а в периферической нервной системе - ганглиями (узлами). Пучки нервных волокон в центральной нервной системе носят названия трактов, в периферической - нервов.
Органы периферической нервной системы
Нервы (нервные стволы) связывают нервные центры головного и спинного мозга с рецепторами и рабочими органами. Они образованы пучками миелиновых и безмиелиновых нервных волокон, которые объединены соединительнотканными компонентами (оболочками): эндоневрием, периневрием и эпиневрием (рис. 114-118). Большинство нервов являются смешанными, т. е. включают афферентные и эфферентные нервные волокна.
Эндоневрий - тонкие прослойки рыхлой волокнистой соединительной ткани с мелкими кровеносными сосудами, окружающие отдельные нервные волокна и связывающие их в единый пучок.
Периневрий - оболочка, покрывающая каждый пучок нервных волокон снаружи и отдающая перегородки вглубь пучка. Он имеет пластинчатое строение и образован концентрическими пластами уплощенных фибробластоподобных клеток, связанных плотными и щелевыми соединениями. Между слоями клеток в пространствах, заполненных жидкостью, располагаются компоненты базальной мембраны и продольно ориентированные коллагеновые волокна.
Эпиневрий - наружная оболочка нерва, связывающая воедино пучки нервных волокон. Он состоит из плотной волокнистой соединительной ткани, содержащей жировые клетки, кровеносные и лимфатические сосуды (см. рис. 114).
Структуры нерва, выявляемые с помощью различных методов окраски. Различные гистологические методы окраски позволяют более детально и избирательно изучить отдельные компоненты
нерва. Так, осмирование дает контрастное окрашивание миелиновых оболочек нервных волокон (позволяя оценить их толщину и дифференцировать миелиновые и безмиелиновые волокна), однако отростки нейронов и соединительнотканные компоненты нерва остаются очень слабо окрашенными или неокрашенными (см. рис. 114 и 115). При окраске гематоксилином-эозином миелиновые оболочки не окрашиваются, отростки нейронов имеют слабо базофильное окрашивание, однако хорошо выявляются ядра нейролеммоцитов в нервных волокнах и все соединительнотканные компоненты нерва (см. рис. 116 и 117). При окраске азотнокислым серебром ярко окрашиваются отростки нейронов; миелиновые оболочки остаются неокрашенными, соединительнотканные компоненты нерва выявляются слабо, их структура не прослеживается (см. рис. 118).
Нервные ганглии (узлы) - структуры, образованные скоплениями нейронов вне ЦНС, - разделяются на чувствительные и автономные (вегетативные). Чувствительные ганглии содержат псевдоуниполярные или биполярные (в спиральном и вестибулярном ганглиях) афферентные нейроны и располагаются преимущественно по ходу задних корешков спинного мозга (чувствительные узлы спинномозговых нервов) и некоторых черепно-мозговых нервов.
Чувствительные ганглии (узлы) спинномозговых нервов имеют веретеновидную форму и покрыты капсулой из плотной волокнистой соединительной ткани. По периферии ганглия находятся плотные скопления тел псевдоуниполярных нейронов, а центральная часть занята их отростками и расположенными между ними тонкими прослойками эндоневрия, несущими сосуды (рис. 121).
Псевдоуниполярные чувствительные нейроны характеризуются сферическим телом и светлым ядром с хорошо заметным ядрышком (рис. 122). Цитоплазма нейронов содержит многочисленные митохондрии, цистерны гранулярной эндоплазматической сети, элементы комплекса Гольджи (см. рис. 101), лизосомы. Каждый нейрон окружен слоем прилежащих к нему уплощенных клеток олигодендроглии или мантийными глиоцитами) с мелкими округлыми ядрами; снаружи глиальной оболочки имеется тонкая соединительнотканная капсула (см. рис. 122). От тела псевдоуниполярного нейрона отходит отросток, разделяющийся Т-образно на периферическую (афферентную, дендритную) и центральную (эфферентную, аксональную) ветви, которые покрываются миелиновыми оболочками. Периферический отросток (афферентная ветвь) заканчивается рецепторами,
центральный отросток (эфферентная ветвь) в составе заднего корешка вступает в спинной мозг (см. рис. 119).
Автономные нервные ганглии образованы скоплениями мультиполярных нейронов, на которых многочисленные синапсы образуют преганглионарные волокна - отростки нейронов, чьи тела лежат в ЦНС (см. рис. 120).
Классификация автономных ганглиев. По локализации: ганглии могут располагаться вдоль позвоночника (паравертебральные ганглии) или впереди него (превертебральные ганглии), а также в стенке органов - сердца, бронхов, пищеварительного тракта, мочевого пузыря и др. (интрамуральные ганглии - см., например, рис. 203, 209, 213, 215) или вблизи их поверхности.
По функциональному признаку автономные нервные ганглии разделяются на симпатические и парасимпатические. Эти ганглии различаются своей локализацией (симпатические лежат пара- и превертебрально, парасимпатические - интрамурально или вблизи органов), а также локализацией нейронов, дающих преганглионарные волокна, характером нейромедиаторов и направленностью реакций, опосредуемых их клетками. Большинство внутренних органов имеют двойную автономную иннервацию. Общий план строения симпатических и парасимпатических нерв ных ганглиев сходен.
Строение автономных ганглиев. Автономный ганглий снаружи покрыт соединительнотканной капсулой и содержит диффузно или группами расположенные тела мультиполярных нейронов, их отростки в виде безмиелиновых или (реже) миелиновых волокон и эндоневрий (рис. 123). Тела нейронов - базофильные, неправильной формы, содержат эксцентрично расположенное ядро; встречаются многоядерные и полиплоидные клетки. Нейроны окружены (обычно не полностью) оболочками из глиальных клеток (сателлитными глиальными клетками, или мантийными глиоцитами). Снаружи от глиальной оболочки располагается тонкая соединительнотканная оболочка (рис. 124).
Интрамуральные ганглии и связанные с ними проводящие пути ввиду их высокой автономии, сложности организации и особенностей медиаторного обмена некоторыми авторами выделяются в самостоятельный метасимпатический отдел автономной нервной системы. В интрамуральных ганглиях описаны нейроны трех типов (см. рис. 120):
) Длинноаксонные эфферентные нейроны (клетки I типа Догеля) с короткими дендритами и длинным аксоном, идущим за пределы узла
к клеткам рабочего органа, на которых он образует двигательные или секреторные окончания.
2) Равноотростчатые афферентные нейроны (клетки II типа Догеля) содержат длинные дендриты и аксон, уходящий за пределы данного ганглия в соседние и образующий синапсы на клетках I и III типов. Входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в центральную нервную систему.
3) Ассоциативные клетки (клетки III типа Догеля) - местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типов. Дендриты этих клеток не выходят за пределы узла, а аксоны направляются в другие узлы, образуя синапсы на клетках I типа.
Рефлекторные дуги в соматическом (анимальном) и автономном (вегетативном) отделах нервной системы обладают рядом особенностей (см. рис. 119 и 120). Основные различия заключаются в ассоциативном и эффекторном звеньях, поскольку рецепторное звено сходно: оно образовано афферентными псевдоуниполярными нейронами, тела которых располагаются в чувствительных ганглиях. Периферические отростки этих клеток образуют чувствительные нервные окончания, а центральные вступают в спинной мозг в составе задних корешков.
Ассоциативное звено в соматической дуге представлено вставочными нейронами, дендриты и тела которых расположены в задних рогах спинного мозга, а аксоны направляются в передние рога, передавая импульсы на тела и дендриты эфферентных нейронов. В автономной дуге дендриты и тела вставочных нейронов расположены в боковых рогах спинного мозга, а аксоны (преганглионарные волокна) покидают спинной мозг в составе передних корешков, направляясь в один из автономных ганглиев, где и оканчиваются на дендритах и телах эфферентных нейронов.
Эффекторное звено в соматической дуге образовано мультиполярными мотонейронами, тела и дендриты которых лежат в передних рогах спинного мозга, а аксоны выходят из спинного мозга в составе передних корешков, направляются к чувствительному ганглию и далее в составе смешанного нерва - к скелетной мышце, на волокнах которой их веточки образуют нейро-мышечные синапсы. В автономной дуге эффекторное звено образовано мультиполярными нейронами, тела которых лежат в составе автономных ганглиев, а аксоны (постганглионарные волокна) в составе нервных стволов и их ветвей направляются к клеткам рабочих органов - гладких мышц, желез, сердца.
Рис. 121. Чувствительный ганглий спинномозгового нерва
Окраска: гематоксилин-эозин
1 - задний корешок; 2 - чувствительный ганглий спинномозгового нерва: 2.1 - соединительнотканная капсула, 2.2 - тела псевдоуниполярных чувствительных нейронов, 2.3 - нервные волокна; 3 - передний корешок; 4 - спинномозговой нерв

Рис. 122. Псевдоуниполярный нейрон чувствительного ганглия спинномозгового нерва и его тканевое микроокружение
Окраска: гематоксилин-эозин
1 - тело псевдоуниполярного чувствительного нейрона: 1.1 - ядро, 1.2 - цитоплазма; 2 - сателлитные глиальные клетки; 3 - соединительнотканная капсула вокруг тела нейрона

Рис. 123. Автономный (вегетативный) ганглий из солнечного сплетения
1 - преганглионарные нервные волокна; 2 - автономный ганглий: 2.1 - соединительнотканная капсула, 2.2 - тела мультиполярных вегетативных нейронов, 2.3 - нервные волокна, 2.4 - кровеносные сосуды; 3 - постганглионарные волокна

Рис. 124. Мультиполярный нейрон автономного ганглия и его тканевое микроокружение
Окраска: железный гематоксилин
1 - тело мультиполярного нейрона: 1.1 - ядро, 1.2 - цитоплазма; 2 - начало отростков; 3 - глиоциты; 4 - соединительнотканная оболочка