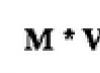У высших растений происходит насасывание воды из почвы корневой системой, проведение ее вместе с растворенными веществами к отдельным органам и клеткам и выведение путем транспирации . В водном обмене у высших растений около 5 % воды используется в ходе фотосинтеза , остальная часть идет на компенсацию испарения и поддержание осмотического давления.
Вода, поступающая из почвы в растения, почти полностью испаряется через поверхность листьев. Это явление называется транспирацией. Транспирация - уникальное явление в наземных экосистемах, играющее важную роль в энергетике экосистем. Рост растений существенно зависит от транспирации. Если влажность воздуха слишком велика, как, например, в тропическом лесу, где относительная влажность приближается к 100 %, то деревья отстают в росте. В этих лесах большая часть растительности представлена эпифитами, по-видимому, из-за отсутствия "транспирационной тяги".
Отношение роста растений (чистой продукции) к количеству транспирированной воды называется эффективностью транспирации . Она выражается в граммах сухого вещества на 1000 г транспирированной воды. Для большинства видов сельскохозяйственных культур и диких видов растений эффективность транспирации равна или менее 2. У засухоустойчивых растений (сорго, просо) она равна 4. У растительности пустынь она не намного больше, так как адаптация у них выражается не в уменьшении транспирации, а в способности прекращать рост при отсутствии воды. В сухой сезон эти растения сбрасывают листья или, как кактусы, закрывают на дневное время устьица.
Растения сухого климата приспосабливается морфологическими изменениями, редукцией вегетативных органов, особенно листьев.
Адаптации животных
|
Животные теряют влагу с испарениями, а также путем выделения конечных продуктов обмена веществ. Компенсацией потерь воды у животных служит ее поступление с пищей и питьем. (н апример, большинство амфибий, некоторые насекомые и клещи). Большая часть животных пустынь никогда не пьет, они удовлетворяют свои потребности за счет воды, поступившей с пищей Другие всасывают ее через покровы тела в жидком или парообразном состоянии . |
|
В неблагоприятных условиях животные часто сами регулируют свое поведение так, чтобы избежать недостатка влаги: переходят в защищенные от иссушения места, ведут ночной образ жизни. Многие животные не покидают пределов переувлажненных местообитаний. Другие животные получает воду в процессе окисления жиров . Например, верблюд, и насекомые - рисовый и амбарный долгоносик и другие. |
Классификация организмов по отношению к влажности среды
Гидатофиты - это водные растения.
Гидрофиты - это растения наземно-водные.
Гигрофиты - наземные растения живущие в условиях повышенной влажности.
Мезофиты - это растения, произрастающие при среднем увлажнении
Ксерофиты - это растения произрастающие с недостаточным увлажнением. Они в свою очередь делятся на:
Суккуленты - сочные растения (кактусы).
Склерофиты - это растения с узкими и мелкими листьями, и свернутыми в трубочки.
Осадки, тесно связанные с влажностью воздуха, представляют собой результат конденсации и кристаллизации водяных паров в высоких слоях атмосферы. В приземном слое воздуха образуются росы, туманы, а при низких температурах наблюдается кристаллизация влаги - выпадает иней.
Одна из основных физиологических функций любого организма - поддержание на достаточном уровне количества воды в теле. В процессе эволюции у организмов сформировались разнообразные приспособления к добыванию и экономному расходованию воды, а также к переживанию засушливого периода. Одни животные пустыни получают воду из пищи, другие за счет окисления своевременно запасенных жиров (например, верблюд, способный путем биологического окисления из 100 г жира получить 107 г метаболической воды); при этом у них минимальна водопроницаемость наружных покровов тела, преимущественно ночной образ жизни и т. д. При периодической засушливости характерно впадание в состояние покоя с минимальной интенсивностью обмена веществ. Наземные растения получают воду главным образом из почвы. Малое количество осадков, быстрый дренаж, интенсивное испарение либо сочетания этих факторов ведут к иссушению, а избыток влаги - к переувлажнению и заболачиванию почв.
Баланс влаги зависит от разницы между количеством выпавших осадков и количеством воды, испарившейся с поверхностей растений и почвы, а также путем транспирации.
4. Влияние концентрации биогенных элементов, солености, рН, газового состава среды, течений и ветера, гравитация, электромагнитных полей на организмы.
Биогенные элементы химические элементы, постоянно входящие в состав организмов и имеющие определённое биологическое значение. Прежде всего это кислород (составляющий 70% массы организмов), углерод (18%), водород (10%), кальций, азот, калий, фосфор, магний, сера, хлор, натрий, железо. Эти элементы входят в состав всех живых организмов, составляют их основную массу и играют большую роль в процессах жизнедеятельности.
Многие элементы имеют большое значение только для определённых групп живых существ (например, бор необходим для растений, ванадий - для асцидий и т.п.). Содержание тех или иных элементов в организмах зависит не только от их видовых особенностей, но и от состава среды, пищи (в частности, для растений - от концентрации и растворимости тех или иных почвенных солей), экологических особенностей организма и других факторов. Элементы, постоянно содержащиеся в организмах млекопитающих, по их изученности и значению можно разделить на 3 группы: элементы, входящие в состав биологически активных соединений (ферменты, гормоны, витамины, пигменты) , они являются незаменимыми; элементы, физиологическая и биохимическая роль которых мало выяснена или неизвестна.
Соленость
Водный обмен теснейшим образом связан с солевым обменом. Он приобретает особое значение для водных организмов (гидробионтов ).
Для всех водных организмов характерно наличие проницаемых для воды покровов тела, поэтому различие в концентрации растворенных в воде солей и солей, определяющих осмотическое давление в клетках организма, ток. создает осмотический Он направлен в сторону большего давления.
У гидробионтов, обитающих в морских и пресноводных экосистемах наблюдаются существенные отличия в адаптациях к концентрации растворенных в водной среде солей.
У большинства морских организмов внутриклеточная концентрация солей близка к таковой в морской воде.
Любые изменения внешней концентрации приводят к пассивному изменению осмотического тока.
Внутриклеточное осмотическое давление меняется соответственно изменению концентрации солей в водной среде. Такие организмы называютпойкилоосмотическими.
К ним относятся все низшие растения (в том числе сине-зеленые водоросли- цианобактерии), большинство морских беспозвоночных животных.
Диапазон толерантности к изменениям концентрации солей у этих организмов невелик; они распространены, как правило, в морских экосистемах с относительно постоянной соленостью .
К другой группе водных организмов относятся так называемые гомойоосмотические.
Они способны активно регулировать осмотическое давление и поддерживать его на определенном уровне независимо от изменений концентрации солей в воде, поэтому их называют также осморегуляторами.
К ним относятся высшие раки, моллюски, водные насекомые. Осмотическое давление внутри их клеток не зависит от химической природы растворенных в цитоплазме солей. Оно обусловлено общим количеством растворенных частиц (ионов). У осморегуляторов активная ионная регуляция обеспечивает относительное постоянство внутренней среды, а также способность избирательно извлекать из воды отдельные ионы и накапливать их в клетках своего организма.
Задачи осморегуляции в пресной воде противоположны таковым в морской.
У пресноводных организмов внутриклеточная концентрация солей всегда выше, чем в окружающей среде.
Осмотический ток всегда направлен внутрь клеток, и эти виды являются гомойосмотическими.
Важным механизмом поддержания у них водно-солевого гомеостаза является активный перенос ионов против градиента концентрации.
У некоторых водных животных этот процесс осуществляется поверхностью тела, но главным местом такого активного транспорта служат специальные образования – жабры.
В ряде случаев покровные образования затрудняют проникновение воды через кожу, например, чешуя, панцири, слизь; тогда активное выведение воды из организма происходит с помощью специализированных органов выделения.
Водно-солевой обмен у рыб представляет собой более сложный процесс, который требует отдельного рассмотрения. Здесь отметим лишь, что он происходит по следующей схеме:
Вода поступает в организм осмотическим путем через жабры и слизистую оболочку желудочно-кишечного тракта, избыток ее выводится через почки. Фильтрационно-реабсорбционная функция почек может меняться в зависимости от соотношения осмотических давлений водной среды и жидкостей организма. Благодаря активному переносу ионов и способности к осморегуляции многие пресноводные организмы, в том числе рыбы, приспособились к жизни в солоноватой и даже в морской воде.
Наземные организмы имеют в той или иной мере специализированные структурно-функциональные образования, обеспечивающие водной-солевой обмен. Известны многочисленные варианты приспособлений к солевому составу среды и его изменениям у обитателей суши. Эти приспособления становятся решающими в тех случаях, когда вода является лимитирующим фактором жизни. Например амфибии , обитают во влажных наземных биотопах благодаря особенностям водно-солевого обмена, которые сходны с обменом у пресноводных животных. По-видимому, такой тип приспособления сохранился в ходе эволюции при переходе из водной среды обитания в наземную.
Для растений аридных (засушливых) зон большое значение в ксерофитных условиях имеет повышенное содержание солей в почве.
Солеустойчивость различных видов растений существенно отличается. На засоленных почвах обитают галофиты – растения, которые переносят большие концентрации солей.
Они накапливают в тканях до 10 % солей, что ведет к повышению осмотического давления и способствует более эффективному насасыванию влаги из засоленных почв.
Некоторые растения выводят избыток солей через специальные образования на поверхности листа, другие обладают способностью связывать соли с органическими веществами.
Реакция среды рН
Распространение и численность организмов существенно зависит от реакции почвы или водной среды.
Загрязнение атмосферного воздуха вследствие сжигания ископаемого топлива (чаще всего диоксидом серы) приводит к отложению сухих ацидогенных частиц и выпадению дождя, состоящего, по сути, из слабой сернистой кислоты. Выпадение таких «кислых дождей» вызывает закисле-ние различных объектов окружающей среды. Сейчас проблема «кислых дождей» стала приобретать глобальный характер.
Влияние закисления сводится к следующему :
Снижение рН ниже 3, также как повышение выше 9, приводит к повреждению протоплазмыкорней большинства сосудистых растений.
Изменение рН в почве вызывает ухудшение условий питания: снижается доступность биогенных элементов для растений.
Снижение рН до 4,0 – 4,5 в почве или донных осадках в водных экосистемах вызывает разложение глинистых пород (алюмосиликатов), вследствие чего среда становится токсичной из-за поступления в воду ионов алюминия (Al).
Железо и марганец, необходимые для нормального роста и развития растений, при низких рН становятся токсичными вследствие перехода в ионную форму.
Пределы устойчивости к закислению почвы у разных растений различны, но только немногие растения могут расти и размножаться при рН ниже 4,5.
При высоких значениях рН, т. е. при подщелачивании, также создаются неблагоприятные условия для жизнедеятельности растений. В щелочных почвах железо, марганец, фосфаты присутствуют в виде малорастворимых соединений и плохо доступны для растений.
Резко отрицательное воздействие оказывает на биоту закисление водных экосистем.Повышенная кислотность действует негативно в трех направлениях:
нарушения осморегуляции, активности ферментов (они имеют оптимумы рН), газообмена;
токсического воздействия ионов металлов;
нарушений в пищевых цепях, изменения пищевого рациона и доступности пищи.
В пресноводных экосистемах определяющую роль в реакции среды играет кальций, который наряду с диоксидом углерода определяет состояние карбонатной системы водных объектов.
Присутствие ионов кальция имеет значение и для поведения остальных компонентов, например железа.
Поступление кальция в воду связано с неорганическим углеродом карбонатных пород, из которых происходит его выщелачивание.
Газовый состав среды обитания
Для многих видов организмов, как бактерий, так и высших животных и растений, концентрация кислорода и двуокиси углерода, которые составляют в атмосферном воздухе 21 % и 0,03 % по объему соответственно, являются лимитирующими факторами.
При этом в наземных экосистемах состав внутренней воздушной среды – атмосферного воздуха – относительно постоянен.
В водных экосистемахколичество и состав газов, растворенных в воде, сильно варьирует.
КИСЛОРОД
В водных объектах – озерах и водохранилищах, богатых органическим веществом – кислород становится фактором, лимитирующим процессы окисления, и тем самым приобретает первостепенную важность.
В воде содержится значительно меньше кислорода, чем в атмосферном воздухе, а вариации его содержания там связаны со значительными колебаниями температуры и растворенных солей.
Растворимость кислорода в воде повышается с понижением температуры и снижается с повышением солености.
Общее количество кислорода в воде обеспечивается поступлением из двух источников:
из атмосферного воздуха (путем диффузии)
из растений (как продукт фотосинтеза).
Физический процесс диффузии из воздуха протекает медленно и зависит от ветра и движения воды.
Поступление кислорода при фотосинтезе определяется интенсивностью процесса диффузии, который зависит, прежде всего, от освещенности и температуры воды.
Вследствие этих причин количество кислорода, растворенного в воде, сильно изменяется в течение суток, в разные сезоны, а также отличается в различных физико-географических и климатических условиях.
УГЛЕКИСЛЫЙ ГАЗ
Диоксид углерода в водных экосистемах не имеет такого большого значения, как кислород.
Растворимость его в воде высокая.
Он образуется в результате дыхания живых организмов, разложения отмерших остатков животных и растений.
Углекислота, образующаяся в воде, вступает в реакцию с известняками, образуя карбонаты и бикарбонаты.
Карбонатная система океанов служит основным резервуаром углекислого газа в биосфере и буфером, поддерживающим концентрацию водородных ионов на уровне, близком к нейтральному.
В целом для всех живых существ кислород и углекислый газ, несомненно, являются лимитирующими факторами существования. Диапазоны величин этих факторов, сложившиеся в ходе эволюции, довольно узки.
Концентрации кислорода, необходимые для дыхания, достаточно постоянны и закрепились в ходе эволюции.
Гомеостаз обеспечивается постоянством параметров внутренней среды организмов; содержание кислорода и углекислого газа в различных тканях и органах поддерживается на относительно постоянном уровне.
Карбонатная система жидкостей организма служит хорошим буфером, обеспечивающим гомеостаз.
течение, ветер
Водные течения :
Глобальные (морские) и локальные.
Глобальные:
Учавствуют в распространении организмов.
Определяют климатические условия многих регионов планеты (гольфстрим)
Локальные:
Влияют на газовый состав среды (воды) (увеличивается концентрация кислорода).
Увеличение течения в водоемах создает увеличение продуктивности сообщества. Стоячая вода создает стрессовые условия, а проточная создает дополнительный источник энергии, повышающий продуктивность.
Способствуют возникновению комплекса морфологических адаптаций, противостоящих течению (?).
Воздушные течения (ветра):
Ветер является лимитирующим фактором, ограничивающим распространение многих животных (насекомые).
Играет важную роль в миграции насекомых. Восходящие токи воздуха подхватывают мелких насекомых на 1-2 км, а затем ветер переносит их на огромные расстояния.
Чем сильнее ветер, тем больше направление миграции совпадает с направлением ветра (бражники, тля и цветочные мухи на Шпицбергене).
Ветер влияет на распределение насекомых по биотопу (поляны, опушки, за кустами, за деревьями ветер слабее).
Определяет возможность полета и активности большинства летающих животных (насекомые, птицы). Активность нападения кровососущих двукрылых.
Влияет на распространение веществ используемых животными в качестве стимуляторов полового поведения (особенно феромоны у насекомых). Запах самки и т.д.
Лимитирует рост растений (в условиях тундры или альпийских лугов растения карликовые). Но влияет и температура.
Определяет особенности миграционного и трофического поведения птиц (парящий полет, миграции мелких птиц).
Сила тяжести
Гравитация влияет на формообразование и физиологию крупных животных (биомеханика). Один из определяющих факторов существования жизни на земле.
Сила тяжести может служить сигнальным фактором у насекомых, в качестве указателя к направлению в открытое пространство. (отрицательный геотропизм ). Стремление вверх по стеблю (против градиента силы тяжести – это стремление к свету, теплу, свободе (особенно для летающих). Эксперименты с голодной саранчой в садках где еда на дне (опустились за едой только через несколько часов).
Положительный геотропизм наблюдается у почвенных животных (Опыты Гилярова с насекомыми в сухой и влажной почвой в садках. Хоть почва и сухая все равно ползли вниз, а там погибали).
Геотропизм может меняться по сезонам в зависимости от условий обитания и зимовки (подкорковые клопы то вниз, то вверх).
ЭЛЕКТОРМАГНИТНЫЕ ПОЛЯ ЗЕМЛИ
1. Многие жужелицы используют магнитное поле землидля ориентации и перемещения в ночное время.
2. Многие ориентируются и передвигаются под углом или параллельно геомагнитным линиям, используя их в ориентации (пчелы, мучные хрущаки, майские жуки.
3. В обычных условиях зрительные и другие ориентиры, а при их отсутствиях включаются магнитные механизмы ориентации.
5. Концепция лимитирующих факторов. "Закон Ю. Либиха". Закон толерантности. Зависимость общего обмена и его интенсивности от массы тела. Правило Аллена, Бергмана, Глогера. Классификация ресурсов. Экологическая ниша. Свойства ниши.
В Мировом океане, к примеру, развитие жизни лимитируется главным образом недостатком азота и фосфора. Поэтому любой подъем на поверхность донных вод, обогащенных этими минеральными элементами, оказывает благотворное влияние на развитие жизни. Особенно ярко это проявляется в тропических и субтропических районах.
закон минимума Ю. Либиха
Живой организм в природных условиях одновременно подвергается воздействия не одного, а многих экологических факторов. Причем любой фактор требуется организму в определенных колическах/дозах. Либих установил, что развитие растения или его состояние зависит не от тех химических эл-в, которые присутствуют в почве в достаточных кол-вах, а от тех, которых не хватает. Если
любого, хотя бы одного из элементов питания в почве меньше, чем требуется данным растениям, то оно будет развиваться ненормально, замедленно, или иметь патологические отклонения.
закон минимума Ю.ЛИБИХА - концепция, согласно которой существование и выносливость организма определяется самым слабым звеном в цепи его экологических потребностей.
Согласно закону минимума жизненные возможности организмов лимитируют те экологические факторы, количество и качество которых близки к необходимому организму или экосистеме минимуму.
Закон толерантности Шелфорда - закон, согласно которому существование вида определяется лимитирующими факторами, находящимися не только в минимуме, но и в максимуме.
Закон толерантности расширяет закон минимума Либиха.
Формулировка
«Лимитирующим фактором процветания организма может быть как минимум, так и максимум экологического влияния, диапазон между которыми определяет степень выносливости (толерантности) организма к данному фактору».
Любой фактор, находящийся в избытке или недостатке, ограничивает рост и развитие организмов и популяций.
Закон толернатности был дополнен в 1975г Ю.Одумом.
Организмы могут иметь широкий диапазон толерантности в отношении одного фактора и узкий диапазон в отношении другого.
Организмы с широким диапазоном толерантности в отношении всех экологических факторов обычно наиболее распространены
Если условия по одному экологическому фактору не оптимальны для вида, то диапазон толерантности может сузиться и в отношении других экологических факторов (например, если содержание азота в почве мало, то требуется больше воды для злаков)
Диапазоны толерантности к отдельным факторам и их комбинациям различны.
Период размножения является критическим для всех организмов, поэтому именно в этот период увеличивается число лимитирующих факторов.
Зависимость общего обмена и его интенсивности от массы тела
Правило Аллена - в экологии - закон, согласно которому выступающие части тела теплокровных животных в холодном климате короче, чем в теплом, поэтому они отдают в окружающую среду меньше тепла. Отчасти правило Аллена справедливо и для побегов высших растений.
Правило Бергмана - в экологии - закон, согласно которому у теплокровных животных, подверженных географической изменчивости, размеры тела особей статистически больше у популяций, живущих в более холодных частях ареала вида.
Правило Глогера - в экологии - закон, согласно которому географические расы животных в теплых и влажных регионах пигментированы сильнее, чем в холодных и сухих регионах. Правило Глогера имеет большое значение в систематике животных.
Ресурсы –количественно выраженные составляющие его жизнедеятельности. Все то что организм потребляет. Ресурсы могут быть органической и неорганической природы (живые и не живые). Доступные и недоступные. Нора, дупло, самка –это все тоже ресурсы. При этом наличный запас всего того что используется организмом и что его окружает постоянно меняется в количественном и качественном отношении. Все это и будет ресурсом.
Ресурсы – вещества из которых состоят тела, энергия, используемая в процессах, места где протекают их стадии жизни. Есть ресурсы пищевые, есть энергетические, пространственные.
Классификация ресурсов (по Тилману -Tilman, 1982):
1.Незаменимые ресурсы
Ни один не в состоянии заменить другой. Скорость роста, которой можно достигнуть при снабжении ресурсом 1 жестко ограничена количеством ресурса 2. Олигофаги.
(-1, +1, 0 – скорость роста биомассы)
2.Взаимозаменяемые ресурсы. Любой из них можно полностью заменить другим. Полифаги. При любой скорости роста количество любого ресурса всегда необходимо. При снижении одного необходимо большее другого и наоборот.
3.Взаимодополняющие (комплементарные) При совместном потреблении организмом данных ресурсов их требуется мньше, чем при раздельном потреблении (для достижения одной и той же скорости роста).
4.Антагонистические. При совместном потреблении скорость роста меньше чем при раздельном потреблении ресурсов. Ядовитые растения в пищу травоядным.
5.Ингибирующие. Это незаменимые ресурсы, но при больших концентрациях являются антагонистами
Введение
1. Среды обитания и экологические факторы
1.1 Воздушная среда
1.2 Водная среда
1.3 Экологические факторы
2. Адаптация
2.1 Адаптация растений к загрязнению атмосферы
2.2 Адаптация растений к засолениям почвы
2.2.1 Растения и тяжелые металлы
2.3 Адаптация растений к биотическим факторам
2.4 Адаптация растений к абиотическим факторам
2.4.1 Влияние температуры
2.4.2 Влияние света на растения
3. Исследовательская часть
Заключение
Использованные информационные ресурсы при выполнении учебно-исследовательской работы
10.Sbio. info Первое био сообщество: информационный портал: [Электрон. ресурс] // Биотические факторы среды и обусловленные ими типы взаимоотношений организмов [сайт] Режим доступа: www.sbio. info/page. php? id=159 (02.04.10)
Приложение
Фото № 1. Листок осины из парка.
Фото №2. Листок, находящийся рядом с проезжей частью.

Фото №3. Пыль на липкой ленте с листа из парка.

Фото №4. Пыль на липкой ленте с листа, находящегося рядом с проезжей частью.

Фото №5. Лишайник на стволе дерева в лесопарке.
Реакции на неблагоприятные факторы среды только при некоторых условиях являются губительными для живых организмов, а в большинстве случаев они имеют адаптивное значение. Поэтому эти ответные реакции были названы Селье «общим адаптационным синдромом». В более поздних работах термины «стресс» и «общий адаптационный синдром» он употреблял как синонимы.
Адаптация — это генетически детерминированный процесс формирования защитных систем, которые обеспечивают повышение устойчивости и протекание онтогенеза в неблагоприятных для него условиях.
Адаптация является одним из важнейших механизмов, который повышает устойчивость биологической системы, в том числе растительного организма, в изменившихся условиях существования. Чем лучше организм адаптирован к какому-то фактору, тем он устойчивее к его колебаниям.
Генотипически обусловленная способность организма изменять метаболизм в определенных пределах в зависимости от действия внешней среды называется нормой реакции . Она контролируется генотипом и свойственна всем живым организмам. Большинство модификаций, которые возникают в пределах нормы реакции, имеют адаптивное значение. Они соответствуют изменениям среды обитания и обеспечивают лучшую выживаемость растений при колебаниях условии окружающей среды. В этой связи такие модификации имеют эволюционное значение. Термин «норма реакции» введен В.Л. Йогансеном (1909).
Чем больше способность вида или сорта модифицироваться в соответствии с окружающей средой, тем шире его норма реакции и выше способность к адаптации. Это свойство отличает устойчивые сорта сельскохозяйственных культур. Как правило, несильные и кратковременные изменения факторов внешней среды не приводят к существенным нарушениям физиологических функций растений. Это обусловлено их способностью сохранять относительное динамическое равновесие внутренней среды и устойчивость основных физиологических функций в условиях изменяющейся внешней среды. В то же время резкие и продолжительные воздействия приводят к нарушению многих функций растения, а нередко и к его гибели.
Адаптация включает в себя все процессы и приспособления (анатомические, морфологические, физиологические, поведенческие и др.), которые способствуют повышению устойчивости и способствуют выживанию вида.
1. Анатомо-морфологические приспособления . У некоторых представителей ксерофитов длина корневой системы достигает несколько десятков метров, что позволяет растению использовать грунтовую воду и не испытывать недостатка влаги в условиях почвенной и атмосферной засухи. У других ксерофитов наличие толстой кутикулы, опушенность листьев, превращение листьев в колючки уменьшают потери воды, что очень важно в условиях недостатка влаги.
Жгучие волоски и колючки защищают растения от поедания животными.
Деревья в тундре или на больших горных высотах имеют вид приземистых стелющихся кустарников, зимой они засыпаются снегом, который защищает их от сильных морозов.
В горных районах с большими суточными колебаниями температуры растения часто имеют форму распластанных подушек с плотно расположенными многочисленными стеблями. Это позволяет сохранять внутри подушек влагу и относительно равномерную в течение суток температуру.
У болотных и водных растений формируется специальная воздухоносная паренхима (аэренхима), которая является резервуаром воздуха и облегчает дыхание частей растения, погруженных в воду.
2. Физиолого-биохимические приспособления . У суккулентов приспособлением для произрастания в условиях пустынь и полупустынь является усвоение СО 2 в ходе фотосинтеза по CAM-пути. У этих растений устьица днем закрыты. Таким образом, растение сохраняет внутренние запасы воды от испарения. В пустынях вода является главным фактором, ограничивающим рост растений. Устьица открываются ночью, и в это время происходит поступление СО 2 в фотосинтезирующие ткани. Последующее вовлечение СО 2 в фотосинтетический цикл происходит днем уже при закрытых устьицах.
К физиолого-биохимическим приспособлениям относятся способность устьиц открываться и закрываться, в зависимости от внешних условий. Синтез в клетках абсцизовой кислоты, пролина, защитных белков, фитоалексинов, фитонцидов, повышение активности ферментов, противодействующих окислительному распаду органических веществ, накопление в клетках сахаров и ряд других изменений в обмене веществ содействует повышению устойчивости растений к неблагоприятным условиям внешней среды.
Одна и та же биохимическая реакция может осуществляться несколькими молекулярными формами одного и того же фермента (изоферментами), при этом каждая изоформа проявляет каталитическую активность в относительно узком диапазоне некоторого параметра окружающей среды, например температуры. Наличие целого ряда изоферментов позволяет растению осуществлять реакцию в значительно более широком диапазоне температур, по сравнению с каждым отдельным изоферментом. Это дает возможность растению успешно выполнять жизненные функции в изменяющихся температурных условиях.
3. Поведенческие приспособления, или избегание действия неблагоприятного фактора . Примером могут служить эфемеры и эфемероиды (мак, звездчатка, крокусы, тюльпаны, подснежники). Они проходят весь цикл своего развития весной за 1,5-2 месяца, еще до наступления жары и засухи. Таким образом, они как бы уходят, или избегают попадания под влияние стрессора. Подобным образом раннеспелые сорта сельскохозяйственных культур формируют урожай до наступления неблагоприятных сезонных явлений: августовских туманов, дождей, заморозков. Поэтому селекция многих сельскохозяйственных культур направлена на создание раннеспелых сортов. Многолетние растения зимуют в виде корневищ и луковиц в почве под снегом, защищающим их от вымерзания.
Адаптация растений к неблагоприятным факторам осуществляется одновременно на многих уровнях регуляции — от отдельной клетки до фитоценоза. Чем выше уровень организации (клетка организм, популяция) тем большее число механизмов одновременно участвует в адаптации растений к стрессам.
Регуляция метаболических и адаптационных процессов внутри клетки осуществляется с помощью систем: метаболической (ферментативной); генетической; мембранной. Эти системы тесно связаны между собой. Так, свойства мембран зависят от генной активности, а дифференциальная активность самих генов находится под контролем мембран. Синтез ферментов и их активность контролируются на генетическом уровне, в то же время ферменты регулируют нуклеиновый обмен в клетке.
На организменном уровне к клеточным механизмам адаптации добавляются новые, отражающие взаимодействие органов. В неблагоприятных условиях растения создают и сохраняют такое количество плодоэлементов, которое в достаточном количестве обеспечено необходимыми веществами, чтобы сформировать полноценные семена. Например, в соцветиях культурных злаков и в кронах плодовых деревьев в неблагоприятных условиях более половины заложившихся завязей могут опасть. Такие изменения основаны на конкурентных отношениях между органами за физиологически активные и питательные вещества.
В условиях стрессов резко ускоряются процессы старения и опадения нижних листьев. При этом нужные растениям вещества перемещаются из них в молодые органы, отвечая стратегии выживания организма. Благодаря реутилизации питательных веществ из нижних листьев сохраняются жизнеспособными более молодые — верхние листья.
Действуют механизмы регенерации утраченных органов. Например, поверхность ранения покрывается вторичной покровной тканью (раневой перидермой), рана на стволе или ветке зарубцовывается наплывами (каллюсами). При утрате верхушечного побега у растений пробуждаются спящие почки и усиленно развиваются боковые побеги. Весеннее восстановление листьев вместо опавших осенью — это также пример естественной регенерации органов. Регенерация как биологическое приспособление, обеспечивающее вегетативное размножение растений отрезками корня, корневища, слоевища, стеблевыми и листовыми черенками, изолированными клетками, отдельными протопластами, имеет большое практическое значение для растениеводства, плодоводства, лесоводства, декоративного садоводства и пр.
В процессах защиты и адаптации на уровне растения участвует и гормональная система. Например, при действии неблагоприятных условий в растении резко возрастает содержание ингибиторов роста: этилена и абсциссой кислоты. Они снижают обмен веществ, тормозят ростовые процессы, ускоряют старение, опадение органов, переход растения в состояние покоя. Торможение функциональной активности в условиях стресса под влиянием ингибиторов роста является характерной для растений реакцией. Одновременно с этим в тканях снижается содержание стимуляторов роста: цитокинина, ауксина и гиббереллинов.
На популяционном уровне присоединяется отбор, который приводит к появлению более приспособленных организмов. Возможность отбора определяется существованием внутрипопуляционной изменчивости устойчивости растений к разным факторам внешней среды. Примером внутрипопуляционной изменчивости по устойчивости может служить недружность появления всходов на засоленной почве и увеличение варьирования сроков прорастания при усилении действия стрессора.
Вид в современном представлении состоит из большого числа биотипов — более мелких экологических единиц, генетически одинаковых, но проявляющих разную устойчивость к факторам внешней среды. В различных условиях не все биотипы одинаково жизненны, и в результате конкуренции остаются лишь те из них, которые наиболее отвечают данным условиям. То есть, устойчивость популяции (сорта) к тому или иному фактору определяется устойчивостью составляющих популяцию организмов. Устойчивые сорта имеют в своем составе набор биотипов, обеспечивающих хорошую продуктивность даже в неблагоприятных условиях.
Вместе с тем, в процессе многолетнего культивирования у сортов изменяется состав и соотношение биотипов в популяции, что отражается на продуктивности и качестве сорта, часто не в лучшую сторону.
Итак, адаптация включает в себя все процессы и приспособления, повышающие устойчивость растений к неблагоприятным условиям среды (анатомические, морфологические, физиологические, биохимические, поведенческие, популяционные и др.)
Но для выбора наиболее эффективного пути адаптации главным является время, в течение которого организм должен приспособиться к новым условиям.
При внезапном действии экстремального фактора ответ не может быть отложен, он должен последовать незамедлительно, чтобы исключить необратимые повреждения растения. При длительных воздействиях небольшой силы адаптационные перестройки происходят постепенно, при этом увеличивается выбор возможных стратегий.
В этой связи различают три главные стратегии адаптации: эволюционные , онтогенетические и срочные . Задача стратегии — эффективное использование имеющихся ресурсов для достижения основной цели — выживания организма в условиях стресса. Стратегия адаптации направлена на поддержание структурной целостности жизненно важных макромолекул и функциональной активности клеточных структур, сохранение систем регуляции жизнедеятельности, обеспечение растений энергией.
Эволюционные, или филогенетические адаптации (филогенез — развитие биологического вида во времени) — это адаптации, возникающие в ходе эволюционного процесса на основе генетических мутаций, отбора и передающиеся по наследству. Они являются наиболее надежными для выживания растений.
У каждого вида растений в процессе эволюции выработались определенные потребности к условиям существования и приспособленность к занимаемой им экологической нише, стойкое приспособление организма к среде обитания. Влаголюбие и теневыносливость, жароустойчивость, холодоустойчивость и другие экологические особенности конкретных видов растений сформировались в результате длительного действия соответствующих условий. Так, теплолюбивые и короткодневные растения характерны для южных широт, менее требовательные к теплу и длиннодневные растения — для северных. Хорошо известны многочисленные эволюционные адаптации к засухе растений-ксерофитов: экономное расходование воды, глубоко залегающая корневая система, сбрасывание листьев и переход в состояние покоя и другие приспособления.
В этой связи сорта сельскохозяйственных растений проявляют устойчивость именно к тем факторам внешней среды, на фоне которых проводится селекция и отбор продуктивных форм. Если отбор проходит в ряде последовательных генераций на фоне постоянного влияния какого-либо неблагоприятного фактора, то устойчивость сорта к нему может быть существенно увеличена. Закономерно, что сорта селекции НИИ сельского хозяйства Юго-Востока (г. Саратов), более устойчивы к засухе, чем сорта, созданные в селекционных центрах Московской области. Таким же путем в экологических зонах с неблагоприятными почвенноклиматическими условиями сформировались устойчивые местные сорта растений, а эндемичные виды растений устойчивы именно к тому стрессору, который выражен в ареале их обитания.
Характеристика устойчивости сортов яровой пшеницы из коллекции Всероссийского института растениеводства (Семенов и др., 2005)
| Сорт | Происхождение | Устойчивость |
| Энита | Подмосковье | Средне засухоустойчивый |
| Саратовская 29 | Саратовская обл. | Засухоустойчивый |
| Комета | Свердловская обл. | Засухоустойчивый |
| Каразино | Бразилия | Кислотоустойчивый |
| Прелюдия | Бразилия | Кислотоустойчивый |
| Колониас | Бразилия | Кислотоустойчивый |
| Тринтани | Бразилия | Кислотоустойчивый |
| ППГ-56 | Казахстан | Солеустойчивый |
| Ошская | Киргизия | Солеустойчивый |
| Сурхак 5688 | Таджикистан | Солеустойчивый |
| Мессель | Норвегия | Соленеустойчивый |
В природной обстановке условия среды обычно изменяются очень быстро, и времени, в течение которого стрессовый фактор достигает повреждающего уровня, недостаточно для формирования эволюционных приспособлений. В этих случаях растения используют не постоянные, а индуцируемые стрессором защитные механизмы, формирование которых генетически предопределено (детерминировано).
Онтогенетические (фенотипические) адаптации не связаны с генетическими мутациями и не передаются по наследству. Формирование такого рода адаптаций требует сравнительно много времени, поэтому их называют долговременными адаптациями. Одним из таких механизмов является способность ряда растений формировать водосберегающий путь фотосинтеза CAM-типа в условиях водного дефицита, вызванного засухой, засолением, действием низких температур и других стрессорами.
Эта адаптация связана с индукцией экспрессии «неактивного» в нормальных условиях гена фосфоенолпируваткарбоксилазы и генов других ферментов CAM-пути усвоения СО 2 , с биосинтезом осмолитов (пролина), с активацией антиоксидантных систем и изменением суточных ритмов устьичных движений. Все это приводит к очень экономному расходованию воды.
У полевых культур, например, у кукурузы, аэренхима в обычных условиях произрастания отсутствует. Но в условиях затопления и недостатка в тканях кислорода в корнях у нее происходит гибель части клеток первичной коры корня и стебля (апоптоз, или программируемая смерть клеток). На их месте образуются полости, по которым кислород из надземной части растения транспортируется в корневую систему. Сигналом для гибели клеток является синтез этилена.
Срочная адаптация происходит при быстрых и интенсивных изменениях условий обитания. В основе ее лежит образование и функционирование шоковых защитных систем. К шоковым защитным системам относятся, например, система белков теплового шока, которая образуется в ответ на быстрое повышение температуры. Эти механизмы обеспечивают кратковременные условия выживания при действии повреждающего фактора и тем самым создают предпосылки для формирования более надежных долговременных специализированных механизмов адаптации. Примером специализированных механизмов адаптации является новообразование антифризных белков при низких температурах или синтез сахаров в процессе перезимовки озимых культур. Вместе с тем, если повреждающее действие фактора превышает защитные и репарационные возможности организма, то неминуемо наступает смерть. В этом случае организм погибает на этапе срочной или на этапе специализированной адаптации в зависимости от интенсивности и продолжительности действия экстремального фактора.
Различают специфические и неспецифические (общие) ответные реакции растений на стрессор.
Неспецифические реакции не зависят от природы действующего фактора. Они одни и те же при действии высокой и низкой температуры, недостатка или избытка влаги, высокой концентрации солей в почве или вредных газов в воздухе. Во всех случаях в клетках растений повышается проницаемость мембран, нарушается дыхание, возрастает гидролитический распад веществ, увеличивается синтез этилена и абсцизовой кислоты, тормозится деление и растяжение клеток.
В таблице представлен комплекс неспецифических изменений, протекающих у растений под влиянием различных факторов внешней среды.
Изменение физиологических параметров у растений под действием стрессовых условий (по Г.В, Удовенко, 1995)
| Параметры | Характер изменения параметров в условиях | |||
| засухи | засоления | высокой температуры | низкой температуры | |
| Концентрация ионов в тканях | Растет | Растет | Растет | Растет |
| Активность воды в клетке | Падает | Падает | Падает | Падает |
| Осмотический потенциал клетки | Растет | Растет | Растет | Растет |
| Водоудерживающая способность | Растет | Растет | Растет | — |
| Водный дефицит | Растет | Растет | Растет | — |
| Проницаемость протоплазмы | Растет | Растет | Растет | — |
| Интенсивность транспирации | Падает | Падает | Растет | Падает |
| Эффективность транспирации | Падает | Падает | Падает | Падает |
| Энергетическая эффективность дыхания | Падает | Падает | Падает | — |
| Интенсивность дыхания | Растет | Растет | Растет | — |
| Фотофосфорилирование | Снижается | Снижается | — | Снижается |
| Стабилизация ядерной ДНК | Растет | Растет | Растет | Растет |
| Функциональнаяя активность ДНК | Снижается | Снижается | Снижается | Снижается |
| Концентрация пролина | Растет | Растет | Растет | — |
| Содержание водорастворимых белков | Растет | Растет | Растет | Растет |
| Синтетические реакции | Подавлены | Подавлены | Подавлены | Подавлены |
| Поглощение ионов корнями | Подавлено | Подавлено | Подавлено | Подавлено |
| Транспорт веществ | Подавлен | Подавлен | Подавлен | Подавлен |
| Концентрация пигментов | Падает | Падает | Падает | Падает |
| Деление клеток | Тормозится | Тормозится | — | — |
| Растяжение клеток | Подавлено | Подавлено | — | — |
| Число плодоэлементов | Снижено | Снижено | Снижено | Снижено |
| Старение органов | Ускорено | Ускорено | Ускорено | — |
| Биологический урожай | Понижен | Понижен | Понижен | Понижен |
Исходя из данных таблицы видно, что устойчивость растений к нескольким факторам сопровождается однонаправленными физиологическими изменениями. Это дает основание считать, что повышение устойчивости растений к одному фактору может сопровождаться повышением устойчивости к другому. Это подтверждено экспериментами.
Опытами в Институте физиологии растений РАН (Вл. В. Кузнецов и др.) показано, что кратковременная тепловая обработка растений хлопчатника сопровождается повышением их устойчивости к последующему засолению. А адаптация растений к засолению приводит к повышению их устойчивости к высокой температуре. Тепловой шок повышает способность растений приспосабливаться к последующей засухе и, наоборот, в процессе засухи повышается устойчивость организма к высокой температуре. Кратковременное воздействие высокой температурой повышает устойчивость к тяжелым металлам и УФ-Б облучению. Предшествующая засуха способствует выживанию растений в условиях засоления или холода.
Процесс повышения устойчивости организма к данному экологическому фактору в результате адаптации к фактору иной природы называется кросс-адаптацией .
Для изучения общих (неспецифических) механизмов устойчивости большой интерес представляет ответ растений на факторы, вызывающие у растений водный дефицит: на засоление, засуху, низкие и высокие температуры и некоторые другие. На уровне целого организма все растения реагируют на водный дефицит одинаково. Характерно угнетение роста побегов, усиление роста корневой системы, синтеза абсцизовой кислоты, снижение устьичной проводимости. Спустя некоторое время, ускоренно стареют нижние листья, и наблюдается их гибель. Все эти реакции направлены на снижение расходования воды за счет сокращения испаряющей поверхности, а также за счет увеличения поглотительной деятельности корня.
Специфические реакции — это реакции на действие какого-либо одного стрессового фактора. Так, фитоалексины (вещества со свойствами антибиотиков) синтезируются в растениях в ответ на контакт с болезнетворными микроорганизмами (патогенами).
Специфичность или не специфичность ответных реакций, подразумевает, с одной стороны, отношение растения к различным стрессорам и, с другой стороны, характерность реакций растений различных видов и сортов на один и тот же стрессор.
Проявление специфических и неспецифических ответных реакций растений зависит от силы стресса и скорости его развития. Специфические ответные реакции возникают чаще, если стресс развивается медленно, и организм успевает перестроиться и приспособиться к нему. Неспецифические реакции обычно возникают при более кратковременном и сильном действии стрессора. Функционирование неспецифических (общих) механизмов устойчивости позволяет растению избегать больших затрат энергии для формирования специализированных (специфических) механизмов адаптации в ответ на любое отклонение от нормы условий их обитания.
Устойчивость растений к стрессовому воздействию зависит от фазы онтогенеза. Наиболее устойчивы растения и органы растений в покоящемся состоянии: в виде семян, луковиц; древесные многолетние — в состоянии глубокого покоя после листопада. Наиболее чувствительны растения в молодом возрасте, так как в условиях стресса процессы роста повреждаются в первую очередь. Вторым критическим периодом является период формирования гамет и оплодотворения. Действие стресса в этот период приводит к снижению репродуктивной функции растений и снижению урожая.
Если стрессовые условия повторяются и имеют небольшую интенсивность, то они способствуют закаливанию растений. На этом основаны методы повышения устойчивости к низким температурам, жаре, засолению, повышенному содержанию в воздухе вредных газов.
Надежность растительного организма определяется его способностью не допускать или ликвидировать сбои на разных уровнях биологической организации: молекулярном, субклеточном, клеточном, тканевом, органном, организменном и популяционном.
Для предотвращения сбоев в жизнедеятельности растений под влиянием неблагоприятных факторов используются принципы избыточности , гетерогенности функционально равнозначных компонентов , системы репарации утраченных структур .
Избыточность структур и функциональных возможностей — один из основных способов обеспечения надежности систем. Избыточность и резервирование имеет многообразные проявления. На субклеточном уровне повышению надежности растительного организма способствуют резервирование и дублирование генетического материала. Это обеспечивается, например, двойной спиралью ДНК, увеличением плоидности. Надежность функционирования растительного организма в изменяющихся условиях поддерживается также благодаря наличию разнообразных молекул информационной РНК и образованию гетерогенных полипептидов. К ним относятся и изоферменты, которые катализируют одну и ту же реакцию, но отличаются по свои физико-химическим свойствам и устойчивостью структуры молекул в изменяющихся условиях среды.
На уровне клетки пример резервирования — избыток клеточных органелл. Так, установлено, что для обеспечения растения продуктами фотосинтеза достаточно части имеющихся хлоропластов. Остальные хлоропласты как бы остаются в резерве. То же касается и общего содержания хлорофилла. Избыточность проявляется также в большом накоплении предшественников для биосинтеза многих соединений.
На организменном уровне принцип избыточности выражается в образовании и в разновременной закладке большего, чем требуется для смены поколений, числа побегов, цветков, колосков, в огромном количестве пыльцы, семязачатков, семян.
На популяционном уровне принцип избыточности проявляется в большом числе особей, различающихся по устойчивости к тому или иному стрессовому фактору.
Системы репарации также работают на разных уровнях — молекулярном, клеточном, организменном, популяционном и биоценотическом. Репаративные процессы идут с затратой энергии и пластических веществ, поэтому репарация возможна только при сохранении достаточной интенсивности обмена веществ. Если обмен веществ прекращается, то прекращается и репарация. В экстремальных условиях внешней среды особенно большое значение имеет сохранение дыхания, так как именно дыхание обеспечивает энергией репарационные процессы.
Восстановительная способность клеток адаптированных организмов определяется устойчивостью их белков к денатурации, а именно устойчивостью связей, которые определяют вторичную, третичную и четвертичную структуру белка. Например, устойчивость зрелых семян к высоким температурам, как правило, связана с тем, что после обезвоживания их белки приобретают устойчивость к денатурации.
Главным источником энергетического материала как субстрата дыхания является фотосинтез, поэтому от устойчивости и способности фотосинтетического аппарата восстанавливаться после повреждений зависит энергообеспечение клетки и связанные с ним репарационные процессы. Для поддержания фотосинтеза в экстремальных условиях в растениях активизируется синтез компонентов мембран тилакоидов, происходит торможение окисления липидов, восстанавливается ультраструктура пластид.
На организменном уровне примером регенерации может служить развитие замещающих побегов, пробуждение спящих почек при повреждении точек роста.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Задание 1. Адаптация растений к распространению семян
Установите, как шла адаптация растений к распространению семян за счет насекомых, птиц, млекопитающих и человека. Заполните таблицу.
Адаптация растений к распространению семян
|
№ п/п |
Виды растений |
Насекомые |
Птицы |
Млеко- питающие |
Человек |
|
культурный | |||||
|
войлочный | |||||
|
трехраздельная | |||||
|
незабудковая | |||||
|
Репейник обыкновенный |
Какими свойствами обладают семена перечисленных в таблице растений, которые способствуют распространению семян найденными Вами способами? Приведите конкретные примеры.
Взаимодействие двух популяций теоретически можно представить в виде парных комбинаций символов «+», «-», «0», где «+» обозначает выгоду для популяции, «-» – ухудшение состояния популяции, то есть вред, и «0» – отсутствие значимых изменений при взаимодействии. Используя предложенную символику, дайте определение типам взаимодействия, приведите примеры взаимоотношений и составьте в тетради таблицу.
Биотические взаимоотношения
|
взаимоотношений |
Символическое обозначение |
Определение взаимоотношений |
Примеры взаимоотношений данного типа |
1. Используя раздаточный дидактический материал, составьте пищевую сеть экосистемы озера.
2. При каких условиях озеро не будет изменяться длительное время?
3. Какие действия людей могут привести к быстрому разрушению озерной экосистемы?
Индивидуальное задание по модулю «От экологии организмов до экологии экосистем» Вариант 6
Задание 1. Адаптация живых организмов к экстремальным условиям жизни
Многие организмы в течение жизни периодически испытывают влияние факторов, сильно отличающихся от оптимума. Им приходится переносить и сильную жару, и морозы, и летние засухи, и пересыхание водоемов, и нехватку пищи. Как приспосабливаются они к таким экстремальным условиям, когда нормальная жизнь сильно затруднена? Приведите примеры основных путей адаптации к перенесению неблагоприятных условий жизни
Задание 2. Биотические взаимоотношения.
Определите по графикам, к каким последствиям могут привести взаимоотношения между двумя близкими, совместно обитающими в одной экологической нише видами организмов? Как называется данное взаимоотношение? Ответ поясните.
Рис.11. Рост численности двух видов инфузорий-туфелек (1 – туфелька хвостатая, 2 – туфелька золотистая):
А – при выращивании в чистых культурах с большим количеством пищи (бактерий); Б – в смешанной культуре, с тем же количеством пищи
Задание 3. Природные экосистемы Южного Урала
1. Составьте пищевую сеть речной экосистемы.
2. При каких условиях река не будет изменяться длительное время?
3. Какие действия людей могут привести к быстрому разрушению речной экосистемы?
4. Охарактеризуйте трофическую структуру экосистемы с помощью экологических пирамид численности, биомасс, энергии.
Очень редко семена прорастают на самом растении, как это наблюдается у так называемых живородящих представителей мангровых лесов. Гораздо чаще семена или плоды с заключенными в них семенами полностью теряют связь с материнским растением и начинают самостоятельную жизнь где-то в другом месте.
Часто семена и плоды падают недалеко от материнского растения и здесь же прорастают, давая начало новым растениям. Но чаще всего животные, ветер или вода уносят их на новые места, где они, если условия окажутся подходящими, могут прорасти. Так происходит расселение - необходимый этап в семенном размножении.
Для обозначения любых частей растения, служащих для расселения, существует очень удобный термин диаспора (от гроч. diaspeiro - рассеиваю, распространяю). Употребляются также такие термины, как «пропагула», «мигрула», «диссеминула» и «гермула», а в русской литературе, кроме того, предложенный В.Н. Хитрово термин «зачаток расселения». В мировой литературе получил распространенно термин «диаспора», хотя он, может быть, и не самый лучший. Основные диаспоры, с которыми мы будем иметь дело в этом разделе, - это семена и плоды, реже - целью соплодия или же, напротив, только части плода, очень редко целое растение.
Первоначально диаспорами цветковых растений являлись отдельные семена. Но, вероятно, уже на ранних этапах эволюции эта функция стала переходить к плодам. У современных цветковых растений диаспорами являются в одних случаях семена (особенно у примитивных групп), в других - плоды. У растений с вскрывающимися плодами, такими, как листовка, боб или коробочка, диаспорой является семя. Но с возникновением сочных плодов (ягод, костянок и пр.), а также нераскрывающихся сухих плодов (орешков, семянок и пр.) диаспорой становится сам плод. В некоторых семействах, например в семействе лютиковых, мы можем наблюдать оба типа диаспор.
У относительно очень небольшого числа цветковых растений диаспоры распространяются без участия каких-либо внешних агентов. Такие растения называются автохорами (от греч. autos - сам и choreo - отхожу, продвигаюсь), а само явленно - автохорией. Но у подавляющего большинства цветковых растений диаспоры распространяются посредством животных, воды, ветра или, наконец, человека. Это аллохоры (от греч. allos - другой).
В зависимости от агента, участвующего в распространении семян и плодов, аллохория подразделяется на зоохорию (от греч. zoon - животное), антропохорию (от греч, anthropos - человек), анемохорию (от греч. anomos - ветер) и гидрохорию (от греч. hydro - вода) (Фёдоров, 1980).
Автохория - распространение семян в результате активности каких-либо структур самого растения либо под влиянием силы тяжести. Например, створки бобов нередко резко скручиваются при вскрывании плода и отбрасывают семена. Опадение диаспор под действием силы тяжести называют барохорией.
Баллистохория - разбрасывание диаспор в результате упругих движений стеблей растений, вызванных порывами ветра, или возникающих, когда ка-кое-либо животное или человек при движении задевает растение. У баллистохорных гвоздичных диаспорами служат семена, а у зонтичных - мерикарпии.
Анемохория - распространение диаспор при помощи ветра. Диаспоры могут при этом распространяться в толще воздуха, по поверхности грунта или воды. Для анемохорных растений адаптивно выгодно увеличение парусности диаспор. Это может достигаться путем уменьшения их размеров. Так, семена Pyroloideae (грушанковых, одно из подсемейств вересковых - Ericaceae ) и орхидных очень мелкие, пылевидные и могут подхватываться даже конвективными потоками воздуха в лесу. Семена грушанковых и орхидных содержат недостаточно питательных веществ для нормального развития проростка. Наличие у этих растений столь мелких семян возможно только потому, что их проростки микотрофны. Другой способ увеличения парусности диаспор - возникновение разнообразных волосков, хохолков, крыльев и т.д. Плоды с крыловидными выростами, которые развиты у ряда древесных растений, в процессе падения с дерева вращаются, что замедляет их падение и позволяет им удаляться от материнского растения. Аэродинамические свойства плода одуванчика и некоторых других, сложноцветных таковы, что позволяют ему подниматься в воздухе под действием ветра благодаря тому, что разросшийся хохолок волосков в форме зонтика отделен от тяжелой содержащей семя тяжелой части семянки, так называемым носиком. Поэтому под действием ветра плод наклоняется, и возникает подъемная сила. Впрочем, у многих других сложноцветных носика нет, а их снабженные волосками плоды также успешно распространяются ветром.
Гидрохория - перенос диаспор с помощью воды. Диаспоры гидрохорных растений имеют приспособления, повышающие их плавучесть и защищающие зародыш от попадания воды.
Зоохория - распространение диаспор животными. Наиболее важные группы животных, распространяющих плоды и семена - птицы, млекопитающие и муравьи. Муравьи обычно разносят односемянные диаспоры или отдельные семена (мирмекохория). Для диаспор мирмекохорных растений характерно наличие элайосом - богатых питательными веществами придатков, которые могут привлекать муравьев также своим внешним видом и запахом. Муравьи не поедают сами семена разносимых диаспор.
Распространение диаспор позвоночными животными можно разделить на три типа. При эндозоохории животные поедают целые диаспоры (обычно сочные) или их части, причем семена проходят через пищеварительный тракт, но не перевариваются там и выводятся наружу. Содержимое семени защищено от переваривания плотной оболочкой. Это может быть спермодерма (в ягодах) или внутренний слой перикарпия (в костянках, пиренариях). Семена некоторых растений не способны к прорастанию до тех пор, пока не пройдут через пищеварительный тракт животного. При синзоохории животные употребляют в пищу непосредственно содержимое семени, богатое питательными веществами. Диаспоры синзоохорных растений обычно окружены достаточно прочной оболочкой (например, орехи), разгрызание которой требует усилий и времени. Некоторые животные создают запасы таких плодов в особых местах или уносят их в гнезда, либо просто предпочитают поедать их вдали от продуцирующего растения. Часть диаспор животные теряют или не используют, что и обеспечивает расселение растения. Эпизоохория - перенос диаспор на поверхности животных. Диаспоры могут иметь выросты, шипы и другие структуры, позволяющие зацепиться за шерсть млекопитающих, перья птиц и т.д. Нередки и клейкие диаспоры.
Под антропохорией понимают распространение диаспор человеком. Хотя большинство растений естественных фитоценозов практически не имеют исторически сложившихся адаптаций к распространению плодов и семян именно человеком, хозяйственная деятельность людей способствовала расширению ареала многих видов. Многие растения были впервые - отчасти преднамеренно, отчасти случайно - завезены на континенты, где они ранее не встречались. Некоторые сорняки по ритму развития и размеру диаспор очень близки к возделываемым растениям, поля которых они засоряют. Это можно рассматривать как адаптацию к антропохории. В результате совершенствования техники земледелия некоторые из таких сорных растений стали очень редкими и заслуживают охраны.
Для некоторых растений характерна гетерокарпия - способность к образованию на одном растении плодов различного строения. Иногда неоднородными оказываются не плоды, а части, на которые распадается плод. Гетерокарпия часто сопровождается гетероспермией - разнокаче-ственностью семян, производимых одним растением. Гетерокарпия и гете-роспермия могут проявляться как в морфологическом, так и анатомическом строении плодов и семян, а также в физиологических особенностях семян. Эти явления имеют важное адаптивное значение. Нередко одна часть производимых растением диаспор имеет приспособления к разносу на далекие расстояния, а другая таких приспособлений не имеет. Первые часто содержат семена, способные к прорастанию на следующий год, а вторые - семена, находящиеся в более глубоком покое и входящие в почвенный банк семян. Гетероспермия и гетерокарпия более обычны у однолетних растений (Тимонин, 2009).